Images Collection

View this article in Search Friendly Plain Text
NOTE: This plain text article interpretation has been digitally created by OCR software to estimate the article text, to help both users and search engines find relevant article content. To read the actual article text, view or download the PDF above.
Environment in Relation to Behavior of Spiders
PETER N. WITT, MD SYRACUSE, NY
Reprinted from the Archives of Environmental Health July 1963,’ Vol. 7, pp. 4-12 Copyright 1963, by American Medical Association
Reprinted from the Archives of Environmental Health July 1963,’ Vol. 7, Pp. 4-12 Copyright 1963, by American Medical Association
1
Environment in
Relation to
PETER N. WITT, MD SYRACUSE, NY
Behavior of Spiders ¡
Introduction Is there any particular reason to choose spiders for the study of the environment in its relation to the behavior of organisms ? By looking over the data which have been obtained in the study of the orb web it may become apparent that this choice has been worthwhile. Everybody has occasionally admired the beautiful spider orbs covered with
Submitted for publication Dec. 27, 1961.
Some of the experiments reported here were supported by a grant from the USPHS.
For more extensive reading on spider webs in general, books like H. C. McCook: American Spiders and Their Spinningwork, published by the author, Philadelphia, 1889; A. Tilquin: La Toile géométrique des Araignées, published by Presses Universitaires, Paris 1942; T. H. Savory: The Spider’s Web, published by F. Warne, London 1952; W. J. Gertsch: American Spiders, published by van Nostrand, New York, 1949, are highly recommended.
glistening drops of dew on a sunny summer morning. Spiders can be made to build such webs in the laboratory. Here each factor in their surroundings can be controlled separately; for instance, the temperature can be changed while light and humidity are kept constant. This permits analysis of the influences which different variables have on one function. The complex web presents a wonderful record of the spider’s behavior as we slowly learn to interpret the significance of web shape and to detect web irregularities. By studying a spider—an animal far removed from the mammal—we may become aware not only of differences, but of some general problems which organisms encounter in trying to survive environmental changes.
The Web.—It is frequently stated that the behavior of invertebrates and spiders in particular is rigid. Innate behavioral patterns
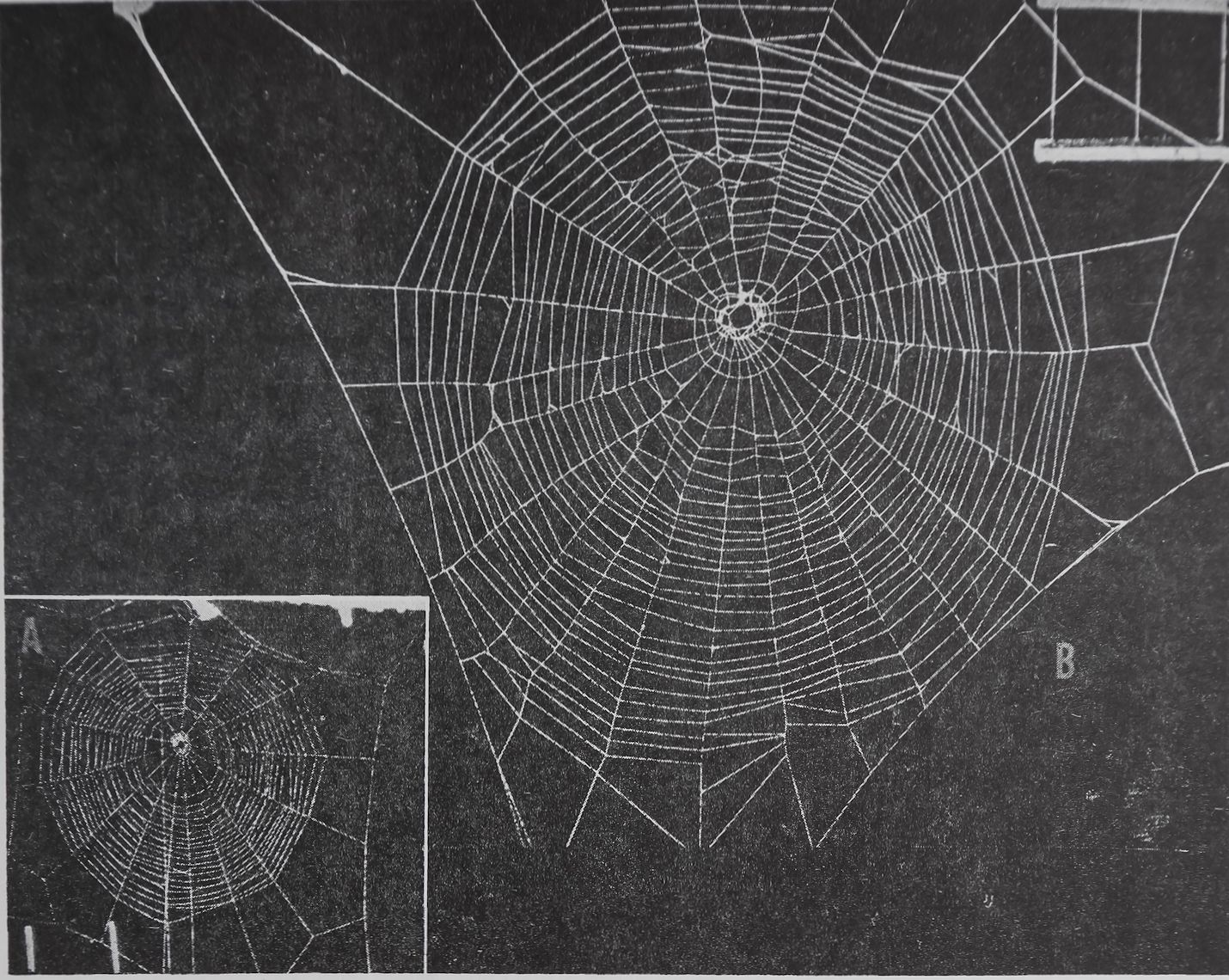
Fig. 1A-—The first web of a baby Araneus diadematus Cl. built 1 week after hatching from the egg. Observe the geometry which has been achieved without previous practice. B, five months later a sister of the spider which built the web in Figure 1A made this web. Both webs are reproduced at the same scale. This web is larger and has a few more radii and spiral turns, but its pattern is basically the same as that in A.
are said to prevail which are independent of experience, require no practice, and are little affected by environment. The building of orb webs by the garden spider Araneus diadematus Cl. (Fig. 3) may serve to evaluate such a statement.
About 2 weeks after hatching from its egg the spiderling builds its first web. At that time most spiders have never seen their parents and have probably never been near another web. But they build their first orbs with a geometrical accuracy and detail indistinguishable from that of an adult spider which has already constructed 100 or more webs during its life 1 (Fig. 1, A and B). It had been suspected that the first 2 weeks might be used by the young spider to practice running and pulling of thread. Mayer2 showed that even if spiderlings were kept in
narrow glass tubes from the first day of their free lives, the webs they built when they were put at large a few weeks later were as flawless as those of their litter-mates. Some changes in web patterns during an individual spider’s lifetime have also been found to be independent of practice: The adult spider of the species Zilla-x-notata Cl. builds an orb web with a free sector, a zone through which a signal thread runs unimpeded by spiral turns. By means of this signal thread the animal in its remote hiding place stays “in touch” with its web (Fig. 2). The young Zilla builds a completely circular web, however. Petrusewiczowa8 prevented young Zilla spiders from building their circular webs by keeping them in narrow glass tubes and feeding them from a pipette. When she let the adults out of their tubes and allowed
11
Witt

6
ARCHIVES OF ENVIRONMENTAL HEALTH

Fig. 2.—A web of an adult spider of the species Zilla-x-notata Cl. In contrast to the webs in Figure 1 the spiral does not cover the whole area. The free sector is divided by the signal thread which leads to the hiding place of the spider.
them to build their first webs she observed normal adult webs with a free sector. Her adult spiders had never built baby webs. This shows that some of the changes in its web which occur during the spider’s life are innate and independent of practice.
There are, however, other changes which depend on the spider’s day-to-day experience. Such small changes affect the rigid basic pattern of the web. The construction of the “normal web” requires an undisturbed functioning of the spider’s body. Muscles, nerves, ganglia, and central regulatory mechanisms must work smoothly so that the movements of the animal’s legs and body can be carried out with the exactness which webbuilding requires. There is some evidence that the spider uses the following procedures to establish the proportions of its geometrical orb: it probes for empty spaces, filling out oversized angles between radii; it turns its

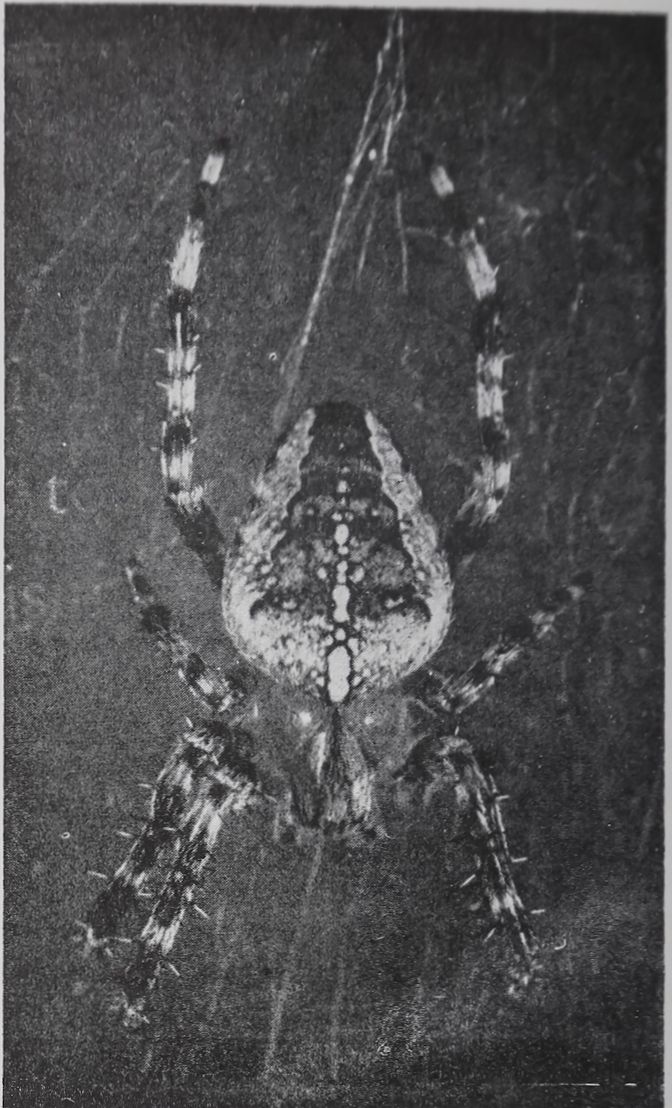
Fig. 3.—An adult spider Araneus diadematus G. in its web. Observe the long legs which contain the lyriform organs and serve in the transmission of haptic signals as auxiliary receptors.
body around a certain angle; it pulls at threads to measure tension and vibration frequency; it estimates distances by spreading its legs or running a definite number of steps,4 and it drops the perpendicular from one point upon a thread to establish the shortest distance.5 It has been shown that a slight disturbance in coordination, central nervous system function, or muscle innervation brought about by relatively low doses of central depressants (pentobarbital sodium6) or stimulants (¿-amphetamine 7) causes distinct and repeatable changes in the web pattern of each individual. Therefore the web-building recorded in the orb web of certain species of spiders is a criterion by which the influence of a number of environmental factors on animals can be analyzed and measured.
Vol 7, July, 1963
BEHAVIOR OF SPIDERS
7
Methods and Materials
Keeping of Spiders.—(For details of method not mentioned here see Witt.0) Unless stated otherwise the species Zilla-x-notata Cl., Araneus diadematus Cl. (Fig. 3), and Araneus sericatus Cl. were used. The animals were caught outdoors or raised from eggs in the laboratory. They were kept indoors in individual wooden or aluminum frames of more than maximal web size, with or without removable glass doors in the front and in the rear. The temperature could be kept constant or programmed to rise and fall in a 24-hour rhythm by means of a Minneapolis Honeywell temperature programmer. Light was turned on and off at predetermined intervals. The animals were fed nearly every day with Drosophila or Musca domestica flies bred in the laboratory, the size and number of flies depending on the size of the spiders. Tap water with or without sugar was administered from the needle of a syringe to the mouth parts of the spider. It took the animals from 1 to 10 minutes to drink a 10 mg. drop.
Procurement of Webs.—Under favorable conditions a web would be built every day. Each web would be made visible by means of ammonia chloride smoke or Krylon white glossy spray paint after the spider had been removed. The white web, if placed against a black ground, could be photographed, together with a scale, on high-contrast 35 mm. copy film. Measurements of web proportions could then be made on the film while the original web was destroyed.
Measurements of Thread Thickness.—In trying to establish the thickness of a thread, microscopic measurements were found unsuitable because threads are not round, and the thickness depends on the tension under which the thread is measured. Calculation of thread thickness as total thread length over the weight of the whole web did not prove practical because of the large experimental error resulting from weighing the light webs. Therefore unsprayed webs were first measured and constituent threads counted against a black ground, and the total thread length was calculated approximately by means of a formula. The webs were then collected on filter paper and digested according to the method of Koch and McMeekin.® After distillation of the ammonia, the nitrogen content could be measured colorimetrically.9 Assuming that the nitrogen content represented total thread protein, the figure for protein per millimeter of thread seemed the most accurate expression for thread thickness.
Measurement of Thread Production.—A number of glands produce the different kinds of thread and glue which spiders use in building their orb webs, hiding places, or cocoons or use for locomotion.10 The silk which forms the skeleton of the orb web (radii, frame, and hub) and is used by spiders in
Witt
locomotion was of interest to us. In order to obtain as much as possible of this thread, its free end was fastened to the axle of a motor which could be turned at variable speed while the spider hung below. The tendency of the spider to let itself down on a thread was used to roll upon the axle just as much silk as the hanging spider would let out. After a long time of such thread-pulling (up to half an hour with 30 and more meters of thread) the spider would drop to the ground without a thread. The silk which had accumulated on the axle was then weighed on a Cahn electromagnetic balance sensitive to 1/ig. Repeated thread-pulling and comparison of figures as well as microscopic inspection of the glands before and after pulling indicated that a high percentage of the stored silk had been depleted. The weight of completed webs of the same spiders, however, was higher. This indicates that the glue and spiral thread, which come from different glands, add considerably to the total web weight. Winding of comparably large quantities from spiders’ abdomens after the cephalothorax had been cut off seemed to demonstrate that no active pressure was necessary for silk production. A phenomenon similar to that described by Morgan and Kwolek11 as “The Nylon Rope Trick” (in which a polyamide cord is pulled from the interface of 2 solutions until 1 solution is used up) might be responsible for thread production.
Results
Light and Temperature.—If we try to understand the world in which a web-building spider lives, we have to exclude most visual impressions. The eyes play such a secondary role in the construction of a geometrical orb web that their removal or covering with black lacquer resulted in webs which were not measurably different from controls.1‘2 Furthermore, most web-building takes place in darkness where very little visual control seems possible. When comparing orb-weaving spiders with jumping spiders Hanstroem 18 found that orb-weavers have primary and secondary visual centers of a much reduced size pointing towards their relative unimportance.
A first experiment seems to indicate that light or its absence does not affect webbuilding as measured by the frequency of new web construction. Two-month-old spiders (Zilla-x-notata Cl.) were kept in glass jars, their webs were destroyed daily, and their web-building was registered 8 A total of
13
ARCHIVES OE ENVIRONMENTAL HEAL’IH
71 animals were kept in constant light and 75 animals in darkness. Both groups were observed in the same room near the open window, where the temperature varied between cool nights and warm days during the summer. To evaluate web-building frequency each spider was observed on 2 consecutive days. An increase from no web on the first day to a web on the second day was counted as +1; a decrease from a web to none as — 1. Building or nonbuilding on both days was rated as zero. The same procedure was used to evaluate spiders under “normal” laboratory conditions. There was no statistically significant difference between the mean frequency of any 2 groups.
A similar experiment was performed with all spiders kept in darkness, but 1 group was kept at an even temperature of 15, 20, or 26 C, while the other went through a temperature change in the daily 24-hour rhythm. No difference of statistical significance in web-building frequency was observed between the 2 groups, nor did the 2 sets differ in the same respect from 68 spiders building under conditions of outdoor temperature and light.
It has been observed that 2 factors may release one action. Noting that there was a slight decrease in web-building under the influence of constant darkness as well as constant temperature, spiders kept in the dark at 15 or 26 C day and night were compared to a group for which temperature and light changed in the 24-hour rhyth^fe of a summer day. The first group, which lived under constant conditions, showed significantly less web-building (P<0.01) than the second. The constant temperature together with the absence of light had resulted in decreasing web-building frequency.6 Thislis still a field open to more detailed study. It was concluded from these experiments as well as from outdoor observations that the 24-hour rhythmic change between light and dark, together with the changing temperature, is recognized by the spider and plays a role in stimulating web-building.
The analysis, however, can be carried further. It can easily be observed and has
been described many times 0,1417 that adult spiders which live together in one room all build their webs simultaneously in the early morning hours just before sunrise. Their web-building time changes with the seasons; in Central Europe and the Northern United States this occurs several hours earlier in summer than in winter. In the laboratory, with a programmed daily rise and fall in temperature and a 12-hour period of light followed by 12 hours of darkness, all spiders would build during the temperature minimum just before the light was turned on in the morning. Spronk15 kept spiders under thermostatically controlled conditions (probably without light changes) and found that almost no spiders built webs at an even temperature and those which did, built at irregular times. He observed, furthermore, that web-building of adult spiders occurred regularly during a period of decreasing temperature or at the early morning temperature minimum. Hardly a web was built while the temperature rose or stayed high.
Wolff and Hempel17 compared meteorological data with the web-building frequency of their spiders. The most important factor in their observations was the equivalent temperature which is defined as 2 times the average temperature of a day plus the mean vapor pressure. Any rise in this figure coincided with a high number of webs on that day in their laboratory; a decrease was accompanied by fewer webs. Only if the equivalent temperature stayed constant during several days, did other factors like changes in barometric pressure seem to influence their web-building figures.
The above evidence points towards a role of temperature and light in the release of the activity of web-building. It will be discussed later in what way such a “releaser” has to meet other conditions to lead to actual web construction. And while the daily temperature changes probably affect higher animals with their inner temperature regulation less than spiders, it is known that the rhythmic alteration between light and darkness influences the activity periods of mice and other mammals too.
Vol 7, July, 1963
14
BEHAVIOR OF SPIDERS
9
In this connection it appears appropriate to mention a book which was published at the end of the 18th century18 which instructed people how to predict the weather from observing the web-building of spiders. In a queer mixture of explanations and observations the author states that if spiders build complete orb webs, a period of dry, sunny weather is sure to follow. In contrast, rain and storm will come when the economical animal does not build at all or even eats its old web. If the building of radii alone during a bad weather period is observed, sun and quiet air can be predicted to follow in 10 to 12 hours, while the building of a complete orb means that the rain will stop even sooner. It seems that this book, which contains much mpre material than can be mentioned here and which is based on observations made during years of the author’s imprisonment, was a great success in its time.
Food.—There is general agreement in the literature that heavy feeding is followed by several days without web-building.16,17,19,20 The interpretation of such behavior runs usually as follows: The spider is no longer hungry and consequently does not need a trap for catching food. Expressing it in a different way we may assume that the hunger drive is too low for releasers like temperature and light to operate. Satiation somehow inhibits web-building.
The effects of food deprivation or hunger are much more complex. Two trends may work in opposite directions: hunger towards a more frequent web-building and larger webs; and the drive to preserve the precious web material (a highly organized protein 21,22) towards economy in web-construction. It has been observed many times that a spider collects and eats its old web. The failure to report whether it had an opportunity to do so in experiments may be responsible for contradictions in the literature regarding web-size during hunger.
Three parameters were observed under different conditions of feeding: the size of the orb webs was measured with a plani-meter on the web photographs, the amount of thread which was produced in the glands
could be established by weighing silk which was rolled up on the axle of a motor, and the body weight of the spiders was regularly checked. It was found that by feeding a spider with a body weight of 138 mg. for 2 weeks on 5 days each week with a 12.5 mg. fly its body weight increased to 159 mg. and that an average of 0.3 mg. of thread per day could be pulled. Though figures for thread production varied widely from day to day, no systematic decrease was observed. We concluded that such a spider showed normal growth and balanced silk production. The average web size of a population of spiders during that period of time increased in young animals and did not change in adults (compare with Reference 1).
The webs of 15 adult spiders and 13 small, young spiders (8-10 mg. body weight) after 1 and 2 days of food deprivation and after daily elimination of webs were compared to webs built by the same spiders previously. No statistically significant changes in web size or web proportions were observed (number of radii, number of spiral turns, diameter), nor was there a measurable change in the amount of thread pulled or in the body weight of the animals.
A group of 9 spiders had their webs eliminated daily and were kept without food for 6 days. Their average web size, showed no significant decrease after that time, but less thread could be pulled. It dropped from an average of 0.49 mg. to 0.39 mg. daily per animal, and body weight decreased by 12%.
After 10 days of food deprivation the same group of spiders on the average built significantly smaller webs of unchanged structure; thread production had further decreased to 0.30 mg. per day per animal, and they had lost an average of 49% in body weight.
Another 17 spiders were kept without food for 20 days. After that period their webs had become significantly smaller and wider-meshed ; they were built with less thread. The body weight of the animals as well as the thread which could be pulled had decreased, but we could still obtain daily 47% of the thread weight which they had produced before deprivation.
15
Witt
10
ARCHIVES OF ENVIRONMENTAL HEALTH
After re feeding had been started, no change in web size or body weight could be observed during the first 3 days. But 10 days’ feeding resulted in a significant increase in the average body weight without changes in web size; the webs stayed small.
In interpreting these preliminary results it appears of interest to note the sequence in which food deprivation and refeeding affected the different parameters. During the first 6 days of hunger, thread production and body weight had decreased, while still largesized webs were built with obviously thinner thread. Even when the animals could no longer produce the full amount of silk, they tried, by building large webs, to catch whatever came near. In later periods of food deprivation webs became smaller and wider-meshed, reflecting the severe decrease in thread synthesis. But some silk production still went on, obviously at the expense of other body constituents as was shown by the surprisingly high loss in body weight. It appears that the production of silk ranges so high in the hierarchy of vital functions in spiders that it is kept up while other tissues shrink. On refeeding, however, thread production or web area did not increase immediately. The increasing body weight with persistently small webs indicates that the spiders first rebuilt their lost tissues. It is also of interest to note in the results of this experiment the close coordination of behavioral (web size) and biochemical (amount of thread) regulatory mechanisms in an emergency situation.
Perception.—Gravity influences the shape of the orb web. In the upper part of the webr angles between radii are larger than at the bottom (Fig. IB), and spiral turns are frequently more widely spaced. In all Araneus webs the vertical diameter of the catching area is longer than the horizontal one. Peters30 performed an experiment which points to the role of gravity during web construction. After the spider in his laboratory had completed the frame, radii, and provisional spiral and had started to build the oval circumference of the sticky spiral, he turned the web 90°. Now the horizontal web di
ameter had become longer than the vertical. The spider corrected this soon by adding additional spiral turns at the new bottom part of the web until the old relationship of a long vertical to a short horizontal diameter was reestablished. Mayer3 shows an orb web which was built under experimental conditions where the frame was laid horizontally. This web possesses a nearly circular shape and an axes ratio of 99.6:100 instead of the normal 62:100 in vertical webs.
Young spiders show, according to Savory,33 negative geotropism. Adult spiders use gravity for orientation in their webs. They went on circuitous ways back to their retreats when the web had been turned while they were in it catching the prey.30
The whole web can also be regarded as part of a sense organ which transmits vibrations. It converts the close sense of touch into a long-distance perception system. While an orb web spider does not identify a fly at close distance if it is placed on a table,34 it can distinguish between different insects which are caught in its web 30 and more centimeters away.
As soon as a tuning fork is held to a spider’s web the animal becomes tense and eventually attacks it.35 In the same manner the spider will attack a drop of glass, bite it, enwrap it, and transport it to its hiding place only because it vibrates.13,36 The drumming of a male on the radii of a female’s web37 serves also as a signal in the elaborate mating pattern.
How closely the spider is guided in open space by its web can be observed in another experiment which is used to catch Zilla for the laboratory. A living fly is thrown into a Zilla web outdoors. The spider rushes along the signal thread to the hub of the web and from there to the prey. While it bites and enwraps its food the signal thread is attached to the inner surface of a small paper bag. When the animal returns from the web along the signal thread it climbs without hesitation into the paper bag and settles down. It will now stay in it even when the bag is carried away, the end of the signal thread being Vol 7, July, 1963 16
BEHAVIOR OF SPIDERS
11
probably the main clue which it uses to recognize the hiding place.
Many reports have appeared in newspapers and magazines about the reaction of spiders to music. A prisoner in his cell is described playing the violin, surrounded by spiders which had appeared from all corners drawn by the music. As soon as he stopped playing, the animals scurry back into their hiding places.’8 Such stories are not as fantastic and unlikely as might be assumed. It can easily be observed that if a spider is approached with a tuning fork it reacts to the airborne sound. It raises its front legs towards the source of the tone.
All such observations found a solid anatomical, physiological, and histological basis through the work of Walcott and Van der Kloot28 and Salpeter and Walcott.29 In the first paper the authors describe the location of an extremely sensitive vibration receptor on the legs of spiders (lyriform organ). With their electrophysiological techniques they were able to define certain properties of the receptor such as its threshold, frequency response, and frequency discrimination. A behavioral procedure, namely the change of stress on the legs, modifies the sensitivity of the receptor thereby probably helping the animal to analyze the intensity of a sound. The second paper establishes the microscopic structure of the organ as a basis for its function.
Such findings together with other observations of spiders’ reactions to sound, enable us to surmise the, world as it appears to the spiders: a world with little light and probably no visual shapes but full of vibrations and surfaces of different tactile texture. The movements and tension of the web’s threads provide the clue by which the animal forms the inner “image” of its varying environment.
Conclusion
After glancing at some aspects of spider behavior as it reflects changes in the surroundings we realize the scarcity of available information. References over 2 centuries were screened,1 but only a small number
17 Witt
were finally quoted. In reviewing the literature and realizing what remains to be done one may conclude with words which Sir Arthur Conan Doyle80 puts in the mouth of Sherlock Holmes as he looks at the evidence in a new case: “We are suffering from a plethora of surmise, conjecture, and hypothesis. The difficulty is to detach the framework of fact . . . from the embellishments of theorists and reporters. Then, having established ourselves on this sound basis, it is our duty to see what inferences may be dravr -and what are the special points upon which the whole mystery turns.”
Peter N. Witt, M.D., State University of New York, Upstate Medical Center, 766 Irving Ave, Syracuse 10, NY.
REFERENCES
1. WTitt, P. N., and Baum, R.: Changes in Orb Webs of Spiders During Growth, Behaviour 16: 309-318, 1960.
2. Mayer, G.: Untersuchungen über Herstellung und Struktur des Radnetzes von Ar anea diadema und Zilla-x-notata mit besonderer Berücksichtigung des Unterschiedes von Jugend- und Altersnetzen, Z. Tierpsychol. 9:337-362, 1953.
3. Petrusewiczowa, E.: Beobachtungen über den Bau des Netzes der Kreuzspinne, Trav. Inst. Biol. f niv. Wilno 13: 1938.
4. Peters, H. M.: Probleme des Kreuzspinnennetzes, Z. Morph, ökol. Tiere 36:179-266, 1939.
5. Witt, P. N.: Ein einfaches Prinzip zur Deutung einiger Proportionen im Spinnennetz, Behaviour 4:172-189, 1952.
6. Witt, P. N.: Die Wirkung von Substanzen auf den Netzbau der Spinne als biologischer Test, Heidelberg, Springer-Verlag, 1956.
7. Witt, P. N.; Brettschneider, L., and Boris, A. P.: Sensitivity to d-Amphetamine in Spiders after Iproniazid and Imipramine, J. Pharmacol. Exp. Ther. 132:183-192, 1961.
8. Koch, F. C, and McMeekin, T. L.: A New DireCt Nesslerization Micro-Kjeldahl Method and a. Modification of the Nessler-Folin Reagent for Ammonia, J. Amer. Chem. Soc. 46:2066-2069, 1924.
9. Christiansen, A.; Baum, R., and Witt, P. N.: Changes in Spider Webs Brought about by Mescaline, Psilocybin and an Increase in Body Weight, J. Pharmacol. Exp. Ther. 136:31-37, 1962.
10. Peters, H. M.: Über den Spinnapparat von Nephila madagascariensis, Z. Naturforsch. 10[B] 10:395-404, 1955.
11. Morgan, P. W., and Kwolek, S. L.: The Nylon Rope Trick, Demonstration of Condensation Polymérization, J. Chem. Educ. 36:182-184, 1959.
ARCHIVES OF ENVIRONMENTAL HEALTH
12. Peters, H. M.: Die Fanghandlung der Kreuzspinne (Experimentelle Analyse des Verhaltens), Z. Vergl. Physiol. 15:693-748, 1931.
13. Hanstroem, B.: Fortgesetzte Untersuchungen über das Araneengehirn, Zool. Jb. Abt. Anat. 59: 455-476, 1934.
14. McCook, H. C.: Hibernation and Winter Habits of Spiders, Proc. Acad. Nat. Sei. Phila. 102-104, 1885.
15. Spronk, F.: Die Abhaengigkeit der Netzbauzeiten der Radnetzspinnen Epeira diademata und Zilla-x-notata von verschiedenen Aussenbedin-gungen, Z. Vergl. Physiol. Abt. C 22:604-613, 1935.
16. Koenig, M.: Beitraege zur Kenntnis des Netzbaus orbiteier Spinnen, Z. Tierpsychol. 8:462-492, 1951.
17. Wolff, D., and Hempel, U.: Versuche über die Beeinflussung des Netzbaues von Zilla-x-notata durch Pervitin, Scopolamin und Strychnin, Z. Vergl. Physiol. 33:497-528, 1951.
18. Quatremere-Disjonval: Araneologie oder
Naturgeschichte der Spinnen nach den neuesten bis jetzt unbekannten Entdeckungen vorzüglich in Rücksicht auf die daraus hergeleitete Angabe atmosphärischer Veränderungen, Translated from the French, Edition 2, Frankfurt am Main, Varren-trapp and Wenner, 1798.
19. Wiehle, H.: Beiträge zur Kenntnis des Radnetzbaues der Epeiriden, Tetragnathiden und Ulo-boriden, Z. Wiss. Biol. Abt. A 8:468-537, 1927.
20. Peters, H. M.: Experimente über die Orien
tierung der Kreuzspinne Epeira diademata im Netz, Zool. Jb. Abt. 411g. Zool. 51:239-259, 1932.
21. Braunitzer, G., and Wolff, D.: Vergleichende chemische Untersuchungen über die Fibroine von Bombyx mori und Ne phila madagascariensis, Z. Naturforschg. [B] 10:404-408, 1955.
22. Fischer, F. G., and Brander, J.: Eine Analyse der Gespinste der Kreuzspinne, Z. Physiol. Chem. 320:92-102, 1960.
23. Savory, T. H.: The Biology of Spiders, London, Sidgwick & Jackson, 1928.
24. Baltzer, F.: Beiträge zur Sinnesphysiologie und Psychologie der Webespinnen, Mitt. Naturforsch. Ges. Bern 165-186, 1923.
25. Boys, C. W.: The Influence of a Tuning-Fork on the Garden Spider, Nature (Lond.) 23: 149-150, 1880.
26. Peters, H. M.: Weitere Untersuchungen über die Fanghandlung der Kreuzspinne, Z. Vergl. Physiol. 19:47-86, 1933.
27. Montgomery, T. H.: Studies on the Habits of Spiders, Particularly Those of the Mating Period, Proc. Acad. Nat. Sei. Phila. 59-149, 1903.
28. Walcott, G, and van der Kloot, W. G.: The Physiology of the Spider Vibration Receptor, J. Exp. Zool. 141:191-244, 1959.
29. Salpeter, M. M., and Walcott, G: An Electron Microscopical Study of a Vibration Receptor in the Spider, Exp. Neurol. 2:232-250, 1960.
30. Conan Doyle, A.: Memoirs of Sherlock Holmes, Dolphin B. Co., Ltd., 1961, p. 10.
Printed and Published in the United Stetes of America
18