Images Collection






View this article in Search Friendly Plain Text
NOTE: This plain text article interpretation has been digitally created by OCR software to estimate the article text, to help both users and search engines find relevant article content. To read the actual article text, view or download the PDF above.
THE FATE OF THE INTACT ORB WEB OF THE SPIDER ARANEUS DIADEMATUS CL.
by
ALAN L. BREED, VICTOR D. LEVINE,
DAVID B. PEAKALL, and PETER N. WITT
(Department of Pharmacology, State University of New York, .. ‘Upstate Medical Center, Syracuse, New York),
Reprinted, from: BEHAVIOUR
Yol. XXIII, 1-2

LEIDEN E. J. BRILL 1964
This is an offprint of an article which has been published originally in BEHAVIOUR. An International Journal of Comparative Ethology, edited by Prof. G. P. Baerends a.o.
This periodical covers the scientific study of animal behaviour and is open to reports of original research in this field, as well as to theoretical papers intended to promote the experimental study of the subject.
The subscription price is D. Gld. 32.— (plus postage) per volume of 4 issues; orders may be placed with your bookseller or directly with the publisher:
E. J. BRILL — PUBLISHER — LEIDEN, The Netherlands
THE FATE OF THE INTACT ORB WEB OF THE SPIDER ARANEUS DIADEMATVS CL. *)
by
ALAN L. BREED 1 2), VICTOR D. LEVINE,
DAVID B. PEAKALL 3), and PETER N. WITT
(Department of Pharmacology, State University of New York, Upstate Medical Center, Syracuse, New York)
(with 6 Figures)
(Rec. 30 – III – 1963)
In the past a method has been developed of testing the effects of drugs by the changes these drugs bring about in spider webs (Witt, 1956). This method is based on the observation that spiders build a new web once a day when the web of the previous day has been destroyed. The new web is similar to the old one being only changed in a characteristic way after a drug has been given. The drug experiments were performed in the laboratory with atmospheric conditions kept,t|imilar to those outdoors on a summer day (Christiansen et al., 1962). It was presumed that if the webs had not been eliminated every day, spiders would have stayed in the same webs several days. Only after the webs have become non-functional they would be rebuilt.
McCook (1889) apparently is of the same opinion when he writes: in reply to Kirby and Spence, “In point of fact the renewal does not take place unless made necessary by the destruction or serious injury of the old snare. The reason assigned for this behavior” … “that the spirals rapidly lose their viscid properties by the action of the air, is not founded on fact as is elsewhere shown. The viscid beads retain their adhesive qualities under ordinary circumstances for a considerable time. It is doubtful if any orb becomes thus disabled in so short a period as that assigned — twenty-four hours — except when exposed to rain.”
1) This work was supported in part by PHS grant B-1794 (C4).
2) Supported as a summer fellow by PHS grant 1GS-102.
3) Supported by general research support grant USPHS 1GS-102.
A. L. BREED, V. D. LEVINE, D. B. PEAKALL & P. N. WITT
There are, however, some indications that this is not so. When the total amount of thread material which went into web-building every day in the laboratory was measured it was found that the Monday webs contained a significantly higher amount of protein than each of the Tuesday to Friday webs. The only difference between these two groups of webs seemed to be that on Saturday and Sunday the old webs had not been destroyed and eliminated. Had the spiders enlarged their Friday webs or had they reinforced the existing threads?
Experiments with food deprived spiders (Witt, 1963) have shown that spinning glands go on producing silk even when no new material is ingested. It could be measured that a spider sooner lost up to 50% of its body weight than stopped thread production. This indicates that the silk gland epithelium goes on producing thread protein even under averse nutritional conditions. It seems difficult to slow down its function. Could the pressure of continuously excreted protein in the lumen of the gland force the animals to spin every day even if the old web is still functional?
On the other hand it seems possible that the filling of the lumen of the silk gland feeds back a signal to the gland epithelium regulating its speed of synthesis of new material. Through such a mechanism a spider would be enabled to slow down protein production until the web becomes non-functional and has to be replaced. After being emptied the gland could speed up production again. What is the latency period of such a mechanism if it exists?
It is one of the purposes of this study to shed some light on the spider’s regulation of production of thread material and to find out whether decreased demand slows down production. This first approach to the problem uses the indirect way of measuring the web built by the intact animal. Except for the studies with radioactively labeled material we did not check the speed of production directly nor did we investigate the gland in isolation. We asked how does the spider behave when it is left undisturbed in its web in the laboratory for one or more days in contrast to the spider whose web is daily eliminated?
An effort was made furthermore to find out how the excess web material was used in the web. It has been shown (Christiansen et al., 1962) that the shorter and thicker thread spun by a heavier spider resulted in a wider meshed web of full size. There are also indications that a spider with less thread material at its disposal after prolonged food deprivation (Witt, 1963) or atropine medication (Witt, 1962) built a wider meshed web. It covered the same area as normal with a shorter thread indicating an interrelationship between amount of thread available and web pattern. Consequently a spider
FATE OF INTACT ORB WEB OF AUANEUS DIADEMATUS
45
with excess thread material could build a web with thicker thread or a narrower meshed web with normal thread thickness or a larger web with a longer thread. A comparison of the proportions in Monday webs with those built on other days of the week with less protein would answer such a question.
According to the segment rule of Peters. (1951) and the presumed mechanism of orientation in spiral construction (Witt, 1952) single web constituents cannot increase or decrease independently. An increase in the number of spiral turns should go together with an increase in web size or an increased number of radii. Comparing proportions in webs built with different amounts of material should support or invalidate these hypotheses.
METHODS
Laboratory raised female Araneus diadematus Cl. were used throughout this experimentation when they became 4 to 5 months old and had moulted 6 to 9 times. Before the experiment began the young spiders were fed about 5 Drosophila flies a week, but as they became larger their diet changed to about 5 house flies (Musca domestica) per week; these spiders also received water daily. During the period of experimentation the spiders were fed one house fly and were given a drop of water from a syringe each day that the webs were destroyed. The days the webs were left intact the spiders were given only a drop of sugar water (about io/xl of a 10% solution) while great care was exercised not to break any fibers while opening and closing the cages. If anything the sugar water on these days would have caused a decrease in web protein since the spider would be called on to use body nitrogen.
The spiders were kept in individual cages with glass windows to observe the web building behavior. These cages are 51 X 51 X 9 cm. made to order by State Metal Awning Co. in Syracuse, New York. On the four sides narrow mesh aluminum screening was put between the frames to allow for air circulation while the front and back panels of the cage were made from double strength window glass in an aluminum frame which could be easily removed or fastened in place with two thumb screws. Most of the scaffold threads of the spider webs were fastened to the screen. However, if a strand was attached to one of the glasses the door was only opened on top and the thread attached with scotch tape to the frame before the whole glass was removed. Care was taken that only minimum distortion of the webs was caused by this procedure.
The experiments were arranged such that the webs were destroyed and removed daily two or more days before the “control day” so that a new
A. L. BREED, V. D. LEVINE, D. B. PEAKALL & P. N. WITT
46
web had to be built with new material every day and there would be no effect of an intact leftover web. After this “control day” the webs were left intact for 1, 2, 3, 4, or 8 days! and three “recovery days” with web removal followed. On each day of the experiment the radii and spiral turns were counted and the horizontal and vertical diameter of the catching area and the free zone were measured as described by Christiansen et al. (1962, figure 1). The total length of the thread was then computed by the formula

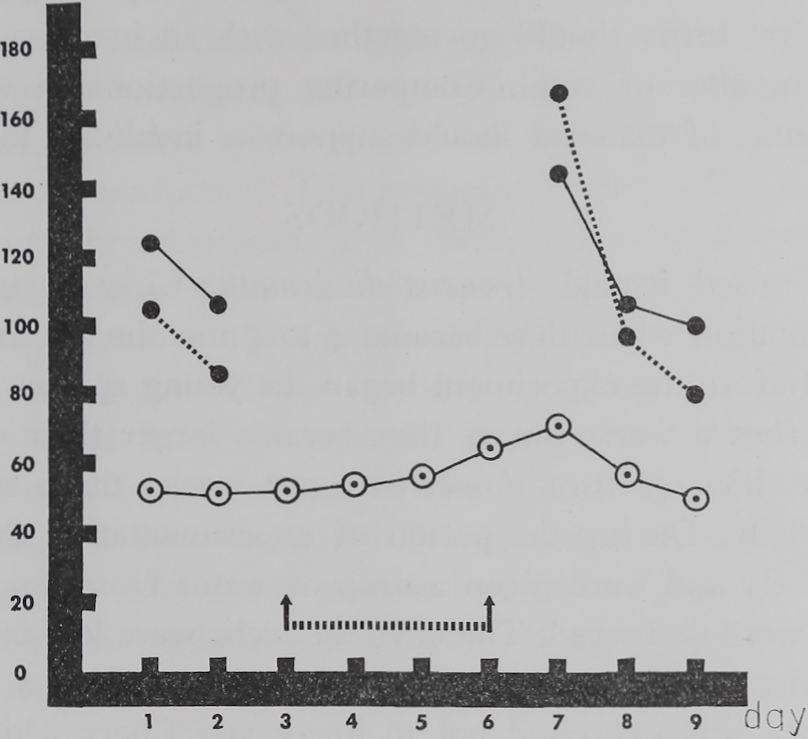
Fig. I. Thread length, web nitrogen and thread thickness before and after a four day period in which webs were not destroyed. E- Each point represents the mean of web measurements of the same 11 spiders during a 9 day observation period (abscissa). Thread length (open circles) is plotted on the ordinate as meters X 3, web nitrogen (full circles connected by dotted line) in micrograms/web, and thread thickness (full circles connected by solid line) as micrograms nitrogen/meter length X 20. The period during which webs were not destroyed (days 3, 4, 5 and 6) is marked by two arrows connected by a broken line. Compare the delayed slight increase in thread length (H” 39%) which could be measured each day in the intact webs and the relatively small increase in thread thickness (+ 36%) with the doubling in nitrogen figures (+94%). According to these measurements the increased amount of web protein on the seventh day — as expressed in the nitrogen figure — was used by the spider to build a longer as well as a thicker thread.
of Dr Walter Baum 1) under the assumption that the spiral consists of evenly spaced perfect circles:
L = nXR + 7r(a + R) where L is the length of the thread of the catching area, n the number of 1
1) We are grateful to Dr W. Baum from Syracuse University for developing the formula for thread length for us.
FATE OF INTACT ORB WEB OF ARANEUS DIADEMATUS
47
radii, R the radius of the catching area (sum of horizontal and vertical diameters divided by 4), k equals the number of spiral turns counted along a radius in the bottom part of the web, and a equals the radius of the inner circular area (free zone).
On the “control day” and the three “recovery days” the webs were removed, digested and analyzed for total nitrogen. For the digestion procedure the catching part of the web was removed from the frame of the web by cutting all the radii except two in the vertical part of the web. A small rectangle of low nitrogen filter paper weighing from 3 to 6 mg was then touched to the lower radius, the latter was cut, the web was rolled onto the filter paper, and all was placed into a tube for digestion.
To the tubes was added 0.2 ml selenium sulfuric acid and digestion proceeded in covered tubes over moderate heat until colorless. Finally 0.4 ml 15 N NaOH, 3 drops of 2% gum ghatti, 3 ml Nessler’s reagent were added and the tubes filled to the 25 ml mark with distilled water. After 10 minutes the light absorption was measured on a Klett spectrophotometer with a green filter three times and the average taken as the indication of total nitrogen. A blank was made in the same way as ¡a sample except only the filter paper was added to be digested. From a standard curve made with ammonium sulfate the results were expressed as gamma nitrogen. This method, a modified P. E. Howe procedure (Howe, 1921) was shown to us by A. Alousi !). It permits an estimate of total nitrogen in a web. If we assume that all threads consist of similar proteins (Braunitzer and Wolff, 1955; Fischer and Brander, i960) the nitrogen figure represents the amount of thread material used to build that web. This has been found to be more accurate than weighing the webs (Christiansen et al., 1962).
On the days when webs were not destroyed the necessary measurements of the dimensions were made without removing the glass panels and precautions were taken to prevent parallax from giving erroneous readings.
For photography of webs the spider was taken out of its cage and kept temporarily in a plastic container. The web was then sprayed with Krylon white glossy spray paint, illuminated with flood lights from the sides, and set against the dark background of an open box which was painted dull black inside. Three vertical white threads 20 mm apart were fastened to the top of the frame (see figure 5a, upper left corner). They appeared on the photograph and permitted enlargement of the negative to the original web size for measurements. Pictures were taken with a Contaflex IV camera on 35 mm high contrast film. 1
1) We are grateful to A. Alousi from this department for showing us the method.
A. L. BREED, V. D. LEVINE, D. B. PEAKALL & P. N. WITT
48
In order to determine whether the web eaten during the night was incorporated into the next web, non-radioactive spiders were placed on radioactive webs. For this experiment two spiders were given C14 labeled alanine and two C14 labeled glucose. The radioactive material was fed to the spiders on Friday and on Monday the animals were removed without damaging the webs. The dimensions of the webs were measured, but it was not possible to determine the radioactivity of this first web. Spiders that had never been used in radioactive experiments were placed carefully on the radioactive webs in the evening. No damage was done to the web and no immediate destruction or eating could be observed. The dimensions of the webs in the cages on the following morning were measured to find out whether webs had been rebuilt. The radioactivity was determined in the digest in a thin-end window gas flow counter against a background of about 20 counts/minute and nitrogen values for these webs were determined as described above. These measurements were repeated on the webs built the following night.
RESULTS
Our experiments show that spiders under laboratory conditions alter their webs even when they are not damaged or destroyed by the experimenters. This is apparent from figures in table I which shoyr a significant increase in spiral turns and table III indicating significant labeling in new webs. It can also be seen from the graphs in figures 2 and 3 which show the significant increase in web nitrogen and thread length on the first morning following the day on which the webs had not been destroyed. When the web of each spider on the first experimental day was compared to its own control is was found that 72% of the 52 spiders observed had different webs after one day. On the following day all but 5 spiders (87%) had webs different from controls. In order to find out whether spiders just enlarged their old webs or doubled the threads or rebuilt the webs completely, direct observation during the night was undertaken. Figures 6a-f show samples of the results. The webs were eliminated by the spiders until only about three radii were left (fig. 6c) and then completely rebuilt in the same manner as new webs (figs 6d-f; Witt, 1961). Spiders cut out sector after sector (figs 6a and b), rolling the threads into a little white ball, and ingested the material. They paused for about 10 minutes after the elimination of each sector, probably in order to liquify the material by preoral digestion. This would make it likely the thread protein was broken down into shorter chains or single amino acids.
Table I and figures 2 and 3 indicate further that the new webs differ from the old ones in a characteristic way. The webs on the first experimental
FATE OF INTACT ORB WEB OF ARANEUS DIADEMATUS
49
day contained a significantly higher amount of nitrogen. If we assume that the nitrogen figure is an indication of the amount of protein, these webs contained significantly more protein. The thread in these webs was longer and was used to produce a spiral with more turns. The control measurements obtained one and two days after the experiments (table I the two bottom lines, figs 2, 3 and 4 the two striped bars on the right) show the
TABLE I
Change of spiral turns, radii, and radius in spider webs which were not
destroyed for one, two, three or four days.
| Days during which web | No. of | % change in No. | % change in | % change in |
| was not destroyed observations | of spiral turns | No. of radii | web radius | |
| One | 48 | + I7-41) | + 6.0 | + 9-3 |
| Two | 34 | + 20.0 0 | + 6.0 | + 9-31) |
| Three | 26 | + 22.4 ó | + 3-9 | 4-ii-2 m |
| Four | 11 | + 34-1 *) | 3-0 | + 1501) |
| Eight | 8 | + 38.2O | + 9-1 | ¡1 1.8 |
| One day after renewed | ||||
| web destruction | 45 | # 2.8 | — 0.2 | + 3-3 |
| Two days after renewed | ||||
| web destruction | 30 | — 2.0 | + 2.2 | |
The mean values for the day preceding the first experimental day are taken as 100%. The relatively high number of observations at the one day values is due to the fact that these values could be obtained from all groups of experiments while 8 day values could be obtained from the last group only.
Observe the lack of change in radius and radii from one to two days in contrast to the change in spiral turns. Later on (3 and 4 days) spirals and radius increased while radii decreased. There was a return close to control day values one and two days after webs had been destroyed again (“Recovery period”)- For further discussion of these figures see text.
measurements of webs which we assume the spiders would have built if the webs had been eliminated on the experimental days (control webs). There would have been no increase in web protein, thread length, and number of spiral turns. It is therefore concluded that non-elimination of webs was the factor which determined the spiders to build new webs which were characteristically different from the old ones.
If webs were not eliminated for more than one day, a further gradual change occurred with time. The amount of protein used for web-building increased from day to day, the second day being significantly higher than the first day (fig. 2). At the end of an 8 day non-elimination period the amount of web protein was more than threefold that of controls (see text
1) Significantly different from controls below the 1% probability level.
A. L. BREED, V. D. LEVINE, D. B. PEAKALL & P. N. WITT
50
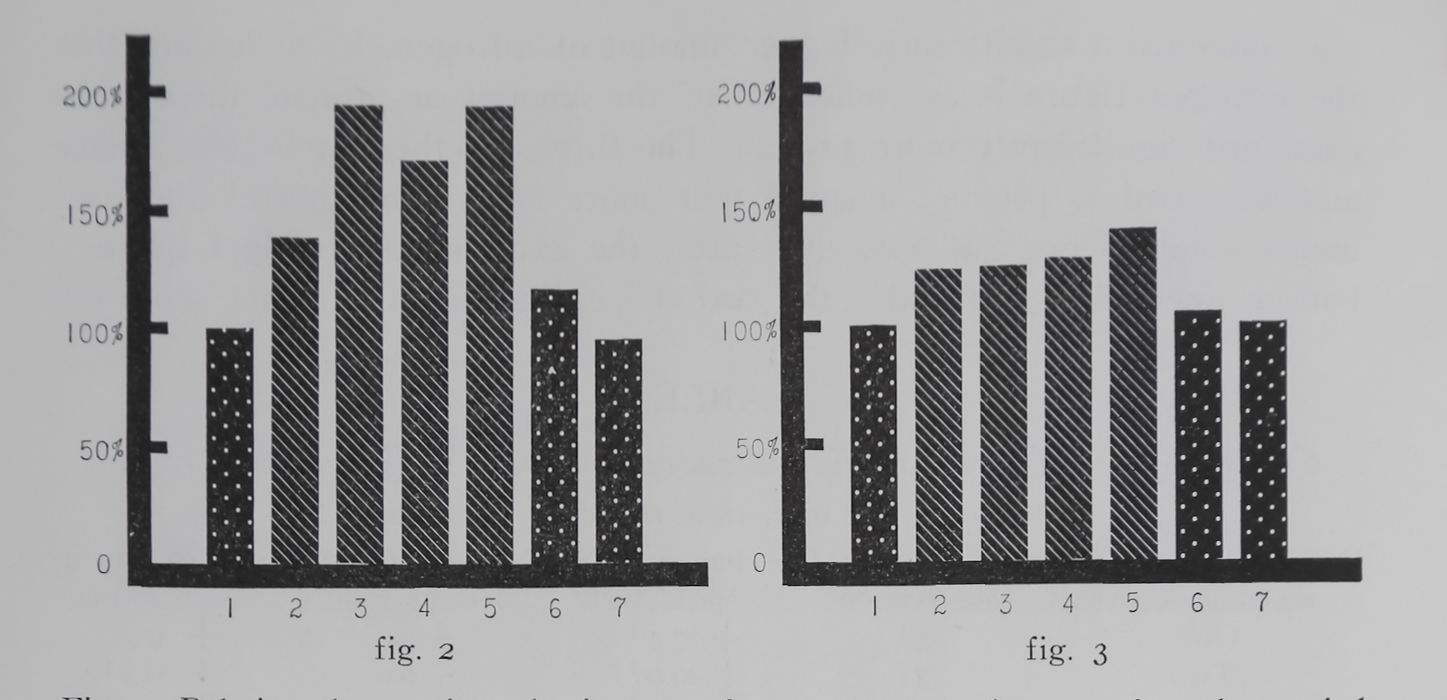
rig. 2. Relative changes in web nitrogen after a one, two, three, or four day period in which webs were not destroyed.^® The dotted bars represent “control days” following web elimination on the preceding day. The first control day (i on the abscissa) shows the mean of 48 web measurements and is arbitrarily set at 100%. The two other control days (6 and 7) follow immediately each period of non-elimination and are derived from 45 and 39 measurements respectively. The first striped bar (2 on the abscissa) stands for the mean of 14 measurements of webs which had not been destroyed for one day; the second (3) for 8 measurements of webs not destroyed for two days; the third (4) for 15 measurements of webs not destroyed for three da:s; the fourth (5) forrai measurements of webs not destroyed for four days. Each experimental group is compared to control webs built by the same spiders. There was a highly significant increase in web nitrogen over controls below the 0.1% probability level on each one of the experimental days. The difference between column 2 and 3 is significant at the 1.8% probability level. After the webs had not been eliminated for 8 days (not represented in this graph) the increase in web nitrogen was 217% over controls (P < 0.001). The results indicate a gradual increase in web protein with increasing length of non-elimination period. Observe also the quick return to control values when webs were eliminated again.
Fig. 3. Relative changes in thread length after a one, two, three, or four day period in which webs were not destroyed. — Abscissa and ordinate as in figure 2; spotted bars controls (#1 on abscissa represents the mean of 48 meaturements, #6 of 45 and #7 of 38 measurements); striped bars: experimental groups (#2 on abscissa is the mean of 48, #3 of 34, #4 of 26, and #5 of 11 measurements). There was a highly significant increase over controls (P < 0.001) at the first experimental day (¡¡I2) and the fourth experimental day (#5). The increase is gradual, #5 being significantly higher than #2 (P < 0.01). Compare the immediate increase in thread length after one day of non-elimination and its further augmentation during the following days with the delayed changes in thread thickness plotted in figure 4.
to fig. 2). It is concluded that these spiders had at the start of their 20 minute web-building period about three times as much fluid protein in the lumen of their silk glands than before the experiment. A look at table I and figures 1, 2, 3, 4, and 5 shows the way in which the excess protein

FATE OF INTACT ORB WEB OF ARANEUS DIADEMATUS
SI
was used. Increasingly longer and thicker threads in quite regular (fig. 5b) larger (up to 4 days) webs were built containing more radii and spiral turns. The increase in web protein cannot be attributed to an increased food intake. On the contrary these spiders were not fed for 4 days during the experiment so that their webs would not be destroyed by flies. They received only sugar water and probably ate their old webs.
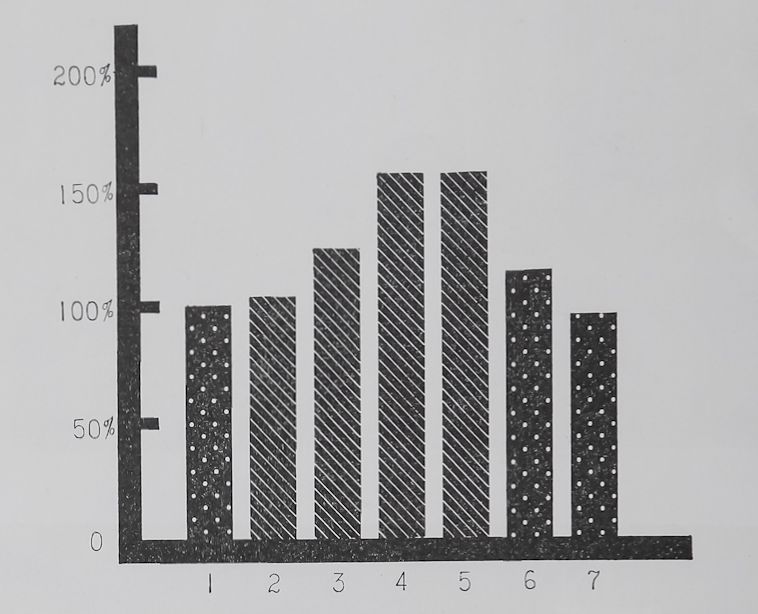
Fig. 4. Relative changes in thread thickness after a one, two, three, or four day period in which webs were not destroyed. — Abscissa and ordinate as in figure 2; each bar represents the mean of the same number of measurements as the corresponding bar in figure 2. There was no statistically significant increase in thread thickness after a one (#2) or two (#3) day period but ä highly significant increase (P < 0.01) after three (#4) and four (#5) days of nón-elimination of webs. After the webs had been left for 8 days the increase in thickness was 140% over control (P < 0.001). A comparison with figure 3 shows that while thread length increased after one day significantly thread thickness took 3 to 4 days to increase significantly. The % increase in thickness during 8 days of the experiment however, was considerably more than the increase in length*. For methods with which these figures were obtained and further discussion of results see text.
In order to avoid fly feeding a separate experiment with fluid food was performed (Metrecal, butterscotch flavor, io/xl per day/spider) during 17 days. In contrast to fly fed controls the Metrecal spiders stopped growing, resuming normal growth only after refeeding with flies was instituted. Not knowing whether the diet would also interfere with web-building, we did not use it further in our experiments.
Our observations which were not extended beyond an 8 day period do not permit a prediction about the upper limit of web growth through non-elimination. Considering the limited capacity of the silk glands no further significant increase in web protein is suspected.
Table I also shows that an increase in the number of spiral turns always goes together with an increase in the number of radii or a larger web area

Fig. 5a and b. Webs built by one spider a before and b after a four day period of non-elimination. Thread length in a 17.9 m» in b 37*7 ni 5 number of spiral turns in a 37, in ft 58; number of radii in a 23, in & 29; area in a 352 cm2, || & 676 cm2. Observe that web b was completely new built using neither the old scaffold threads of a nor its spiral though a had never been eliminated. Both webs were probably built in the same amount of time but b has a considerably longer thread covering a larger area than before. The two photographs illustrate the change in measurements which are shown for a large population of spiders in figures 1-4 and table I.

FATE OF INTACT ORB WEB OF ARANEUS DIADEMATUS
S3
Fig. 6a, b, c, d, e, f. Different stages of spiders’ destruction and reconstruction of webs which were left intact. — In watching spiders during the night it was observed that they cut and ingested sectors of their webs (a and b) until only a few scaffold strands were left (c). Then the spiders began to build new radii (d) followed by a provisional spiral (e) and the sticky or catching spiral (f). The stages of web-building shown in the photographs c-f are like those observed during ordinary fresh web construction (Witt, 1961). The photographs were obtained by interrupting different animals at different stages of reconstruction in the early morning hours around sunrise.

54
A. L. BREED, V. D. LEVINE, D. B. PEAKALL & P. N. WITT
Fig. 6c and d. Explanation see p. 53.



56 A. L. BREED, V. D. LEVINE, D. B. PEAKALL & P. N. WITT
(expressed as a longer web radius). There is consequently no evidence that the spider can build narrower spirals on an unchanged scaffold, — it probably has to change both together. Thread length and thread thickness, however, can change independently as is shown in figures 3 and 4. While thread length increased alone on the first experimental day, thread thickness and length increased both from the third day on.
It is of interest to look for a source for the large amount of thread material on the last experimental days. The body weight of each spider was checked before and after each experiment. In spite of the fact that the spiders received only sugar water during the shorter experiments and sugar water plus 1 fly during the eighth day experiment the animals gained weight. The average weight of 8 spiders was 261 mg. before and 305 mg. after the eight day experiment.
A comparison of the webs of 14 spiders of which the webs had been eliminated the previous day with 11 spiders of which the webs had been destroyed but the threads left for the spiders to eat was supposed to answer the question whether the web material alone or the whole structure of the web constitute the signal for the spider to build a larger web. The 14 spiders which had not had a chance to eat their old web material built webs with 31% less nitrogen the following day (54.1 as compared to 78.2 microgram nitrogen). The 11 controls also decreased nitrogen in the web (78.0 as compared to 98.2 microgram nitrogen) but only by 20%. Though this difference is not significant it seems to indicate that eating the web material is of some importance.
TABLE II
Measurements of web proportions in the experiment with labeled webs before and after the second spider had been introduced
| Day | No. of radii | No. of spiral turns | Vertical diameter mm | Horizontal diameter mm |
| First | 21 | 37 | 103 | hi |
| Second | 20 | 25 | 103 | 120 |
| First | 21 | 26 | 191 | 152 |
| Second | 21 | 18 | 154 | 138 |
| First | 17 | 26 | 173 | 148 |
| Second | 20 | 33 | 234 | 220 |
Each value represents one spider web. The number of radii remained relatively unchanged, but new radii must have been built because the old ones could not be made longer to fit the increased web diameter. The figures indicate an extensive or complete rebuilding of the webs on the second day. A comparison with the figures in table III therefore makes it unlikely that radioactivity on the second day is due to leftover strands but must derive from incorporated material of the first web.
FATE OF INTACT ORB WEB OF ARANEUS DIADEMATUS
57
The figures in table II cannot be explained on the basis of leftover strands of the first radioactive webs. Specifically the change in some proportions without others indicate that webs were completely rebuilt. The percentage of radioactive material incorporated (table III) is similar to that found
TABLE III
Radioactivity of webs produced by 4 “cold” spiders one half day and by the same spiders one and a half days after they had been placed on C14
labeled webs.
| Material fed | Probable | First Day | Probab. | Second Day | Probab. | ||
| to first | activity of | Counts/min | gamma | incorp. | Counts/min | gamma | incorp. |
| spider | original web | nitrogen | %4) | nitrogen | %!) | ||
| C14 alanine | 600- 1,000 | 124 | S4 | 12-20 | Í«f3° | 48 | 13-21 |
| 20/¿g/spider | 114 | 42 | 11-19 | 106 | 38 | 10-18 | |
| C14 glucose | 10-15,000 | 2794 | 72 | 18-28 | 2240 | 52 | 1S-22 |
| 40/ig/spider | 1987 | B | 13-20 | %Mweb | |||
The figures indicate that iájfti to 28% of the web proteins of the old webs had been incorporated into each of the new webs. Such an amount far exceeds the radioactivity of a few strands which might have béeHleft intact from the first web. Material from the old web was most probably reexcreted.
1) Because the actual activity of the first web could not be measured, these values are based on figures obtained under similar conditions with the same dose in other experiments.
when C14 alanine was given to spiders on the evening before web building. Thus it seems certain that the activity is due to new material in the second web and that the substance of the old (radioactive) web is quickly incorporated into the protein with which the web is built.
DISCUSSION
Contrary to McCook’s and our expectations the spider Araneus diadematus Cl. leaves its intact webs in the laboratory only for one or two days. The animals then destroy the webs and ingest them. Such a behavior appears to be rather uneconomical if we assume that the older webs still have their full fly-catching function. The sticky droplets on the spiral, however, may have dried up after one day or the scaffold and the spiral thread has become brittle. Such changes could be measured.
But it was much more surprising to find that the new webs which re-
Bchaviour XXIII
5
SS A. L. BREED, V. D. LEVINE, D. B. PEAKALL & P. N. WITT
placed the eaten ones were different. The main change was the increase in the amount of material which was used for web-building. It was accompanied by changes in web proportions. And this increase occurred day after day over an observation period of 8 days. It means that on each morning following non-destruction of the old web a bigger web was built. The spider apparently behaves differently depending on the old web being destroyed and eliminated or eaten.
There exists the possibility that the difference is due to the material of the old web being immediately incorporated and added to the material for the new web. This seemed at first improbable. There would be about a one hour period in which the old web material is liquified and broken down preorally, ingested, and absorbe^ from the gut, and reexcreted into the lumen of the spinning glan* The experiments with labeled webs tested whether this can be done at such speed. Our results clearly show that fast incorporation takes place. Future experiments have to show whether this is the usual speed of incorporation or whether old web material is faster reexcreted than other food,
Apparently the non-elimination of webs constitutes a signal for spiders to build larger webs the following day. Such a signal could depend on the intact old web or only on the ingestion of the old material. The old material may contain in addition to the bulk of protein a minute amount of a gland stimulating substance. The experiments with destroyed webs which were not eliminated versus eliminated webs seem to indicate that the spider uses both clues. While eating the old web material alone led to a bigger web, it did not cause webs which were as large as after non-elimination. We are inclined to assume that the structure of the intact web as well as eating the material of the old web together form a signal for the spider which regulates size and weight of the new web. This would constitute an example of the interrelationship of a behavioral and a biochemical reaction.
According to Peters’ segment law (Peters, 1951) the number of spiral turns for a given web size depends on the number of radii, or: the steepness with which the spiral approaches the center depends on the size of the angles between radii. Such a relationship between web proportions was explained by Witt (1954) on the basis of the spider’s orientation while it built a web. The small spider while moving at the periphery of the web supposedly uses the shortest way from radius to radius while it constructs the spiral. Such behavior would explain the segment law and is economical. The results of the experiments presented here make such a hypothesis not unlikely. The figures indicate that either the number of radii and the number of spiral turns at a given web size changed together, or with constant radii
FATE OF INTACT ORB WEB OF ARANEUS DIADEMATUS
59
the number of spiral turns and the web size increased together. There is one exception in table I from one to two days. Additional measurements show that the increase of spiral turns without simultaneous increase in radius and radii is caused by a decrease in the diameter of the central free zone. The spider built the spiral more towards the middle. Similar changes have been observed previously after drug application (Witt, 1956).
All our experiments were performed in the laboratory where spiders were confined to individual cages and kept under a controlled diet. It may be asked whether the results are all applicable to outdoor conditions. It appears to us that the principal difference between outdoors and the laboratory lies in the greater possibility for the spider to move around and in a decreased protection of the outdoor webs from dust and larger animals. Diet and atmospheric conditions are probably similar. From the fact that one can see frequently abandoned spider webs in nature it can be concluded that spiders there leave their old webs and build new ones. They could not do that in our boxes where they were forced to create new space by destroying the old web. Consequently our results do not permit us to say whether spiders eat their webs outdoors too. But the protection from wind and other destructive environmental influences should preserve the laboratory webs better, making them functional even longer indoors. One could therefore expect that webs of Araneus diadewiatus Cl. outdoors are renewed even more frequently. Observations are still lacking.
Can we guess at some of the reasons for which the probably economical spider removes its web so frequently? In addition to a quick deterioration of the elastic properties of the silk there may be lack of regulation in the silk glands. If the gland epithelium works at a fixed speed independent of experience the spider will be forced to empty the lumen frequently. And it seems unlikely that such emptying would occur without the building of a new web. The old web is probably ingested to serve as an easily available nitrogen and amino acid source. Future experiments can test such assumptions.
SUMMARY
When spider webs were left intact in the aluminum-glass boxes in the laboratory, most animals ate old webs daily and built new ones. The new webs were built with a larger amount of thread protein, increasing steadily in weight with increasing length of the non-elimination period, to more than 3 times in 8 days. The increased amount of thread material was first used to build larger webs with a longer thread, and later larger webs with thicker threads. A change in the number of spiral turns always went together with a change in web size and/or the number of radii thus confirming Peter’s segment rule. Experiments with radioactive labeled webs indicated that some material for the new web came probably through ingestion and reexcretion of the old web’s
BREED, C.S., FATE OF INTACT ORB WEB OF ARANEUS DIADEMATUS
00
protein. But experiments with intact versus destroyed and non-eliminated webs showed that probably not only old web material but also the intact web structure were clues for larger new webs. The deterioration of silk as well as continuous new thread synthesis in the spinning glands are suspected as reasons for the frequent rebuilding of webs.
REFERENCES
Braunitzer, G. and Wolff, D. (1955). Vergleichende chemische Untersuchungen ueber die Fibroine von Bombyx mori und Nephila madagaskariensis. — Zschr. Natur-forschg. iob, p. 404.
Christiansen, A., Baum, R. and Witt, P. N. (1962). Changes in spider webs brought about by mescaline, psilocybin and an increase in body weight. — J. Pharmacol. 136, ü i|
Fischer, F. G. and Brander, J. (i960). Eine Analyse der Gespinste der Kreuzspinne. — Chemie 320, p. 92.
Howe, P. E. (1921). The determination of proteins in blood |¡§ a micro method. — J. Biol. Chem. 49, p. 109.
Peters, H. M. (1951). Untersuchungen über die Proportionierung im Spinnen Netz.
Dp|- Zschr. Naturforschg. 6b, p. 90.
McCook, H. C. (1889). American Spiders and their Spinningwork. JBfiPhiladelphia, vol. 1, p. 235.
Witt, P. N. (1952). Ein einfaches Prinzip zur Deutung einiger Proportionen im Spinnennetz. = Behaviour, 4, p. 172.
— (1956). Die Wirkung von Substanzen auf den Netzbau der Spinne als biologischer Test. — Heidelberg.
– (1961).Web-Building. 9 Encyclopedia of Biological Sciences, New York, p. 1072.
– (1962). Effects of atropine on spiders’ web building behavior and thread production. — Fed. Proc. 21, p. 180.
— (1963). Environment in relation to the behavior of spiders. — AMA Arch. Envir.
Health 7, p. To.
ZUSAMMENFASSUNG
Liess man Spinnen in Aluminium-Glaskästen ihre Netze, dann verzehrten die meisten nachts die alten Netze und spannen neue. Diese bestanden aus eiweissreicheren Fäden und wurden umso schwerer, je länger das alte Netz unversehrt blieb: nach 8 Tagen war das neue Netz reichlich dreimal so schwer wie das alte. Anfangs wurde der vermehrte Spinnstoff zu grösseren Netzen mit längerem Faden verarbeitet, später zugleich auch mit dickerem. Je grösser das Netz, umso mehr Umgänge und/oder zahlreichere Radien hatte es, gemäss Peters’ Segmentregel. Wenn die Spinnen radioaktiv markierte Netze verzehrt hatten, war das neue Netz am nächsten Morgen auch radioaktiv: wahrscheinlich wird das gefressene Material verdaut und dabei gewonnenes Eiweiss mitversponnen. Da aber Spinnen, die ihre zerstörten alten Netze auf essen durften, kleinere neue Netze bauten als solche, deren alte Netze unversehrt geblieben waren, dürfte nicht nur das Material des alten Netzes, sondern auch das Vorhandensein des unversehrten Netzes Anreiz zum Spinnen des grösseren neuen bilden. Ausser dem schnellen Altern der Seide fordert wohl auch die offenbar ununterbrochene Tätigkeit der den Spinnstoff abscheidenden Drüsen einen so häufigen Netzersatz.