Images Collection










View this article in Search Friendly Plain Text
NOTE: This plain text article interpretation has been digitally created by OCR software to estimate the article text, to help both users and search engines find relevant article content. To read the actual article text, view or download the PDF above.
THE MEASURING FUNCTION OF THE FIRST LEGS OF ARANEUS DIADEMATUS CL.
by
CHARLES F. REED, PETER N. WITT, and ROBERT L. JONES
(Depts. of Psychiatry and Pharmacology, State University of New York, Upstate Medical Center, and IBM Corp., Syracuse, N.Y., U.S.A.)
Reprinted from: BEHAVIOUR Yol. XXV, 1-2
LEIDEN E. J. BRILL
1965
This is an offprint of an article which has been published originally in BEHAVIOUR. An International Journal of Comparative Ethology, edited by Prof. G. P. Baekends a.o.
This periodical covers the scientific study of animal behaviour and is open to reports of original research in this field, as well as to theoretical papers intended to promote the experimental study of the subject.
The subscription price is D. Gld. 32.— (plus postage) per volume of 4 issues; orders may be placed with your bookseller or directly with the publisher:
E. J. BRILL — PUBLISHER — LEIDEN, The Netherlands
THE MEASURING FUNCTION OF THE FIRST LEGS OF ARANEUS DIADEMATUS CL,
by
CHARLES F. REED, PETER N. WITT, and ROBERT L. JONESx) 1 2)
(Depts. of Psychiatry and Pharmacology, State University of New York, Upstate Medical Center, and IBM Corp., Syracuse, N.Y., U.S.A.)
(With 8 Figures)
(Rec. 10-VIII-1964)
In their webs, the orb-weaving spiders leave an evidence of their sensory-motor efficiency which is both intricate and susceptible to objective analysis. The web pattern, whatever else it may be, is an elaborate record of a significant portion of the spider’s behavior, dependent upon the interaction of several more or less independent functions.
A failure in one or more of these functions could have at least two consequences. The geometry of the finished web might be discemably altered, the spider building a web deficient in some aspects. On the other hand, it is conceivable that there are compensatory adaptive alternatives which would permit an unchanged web to be built immediately or after some time.
Such failures in function have been produced experimentally, for example by lesions in the central nervous system of the spider (Witt et al., 1964). Analysis of the web architecture before and after such treatment provides means for relating behavioral changes to sites in the nervous system. There is evidence at hand that the method is capable of considerable subtlety. The web pattern has been shown to be sensitive to small quantities of drugs: the web is altered in particular ways, depending upon the nature and quantity of the substance (Witt, 1958).
The prohibitive amount of time and labor required to measure and compare individual webs has restricted the analysis of the web as a behavioral product. There are variations of a day-to-day nature throughout the life span of the spider (Witt & Baum, i960) which must be taken into account in the assessment of an experimental effect. The measurement of more than one control web per animal is desirable. The variation requires extensive statistical
1) This work was supported in part by USPHS grant B-i794(C4).
2) The technical assistance of Mrs. M. Agi is gratefully acknowledged.
MEASURING FUNCTION OF FIRST LEGS OF ARANEUS
99
treatment. Moreover, the web pattern may be examined on any number of aspects, e.g. placement of radii, relative size of various zones of the web, spiral characteristics. It is one of the objectives of this report to demonstrate the application of modern computer methods to web pattern analysis.
It is essential to the understanding of alterations in web pattern to examine the role of the spider’s legs in the placement of thread. Detailed observation has revealed movements of the first legs which might be interpreted as serving a measuring function. Before discussing those observations it will be necessary to briefly examine a normal web.

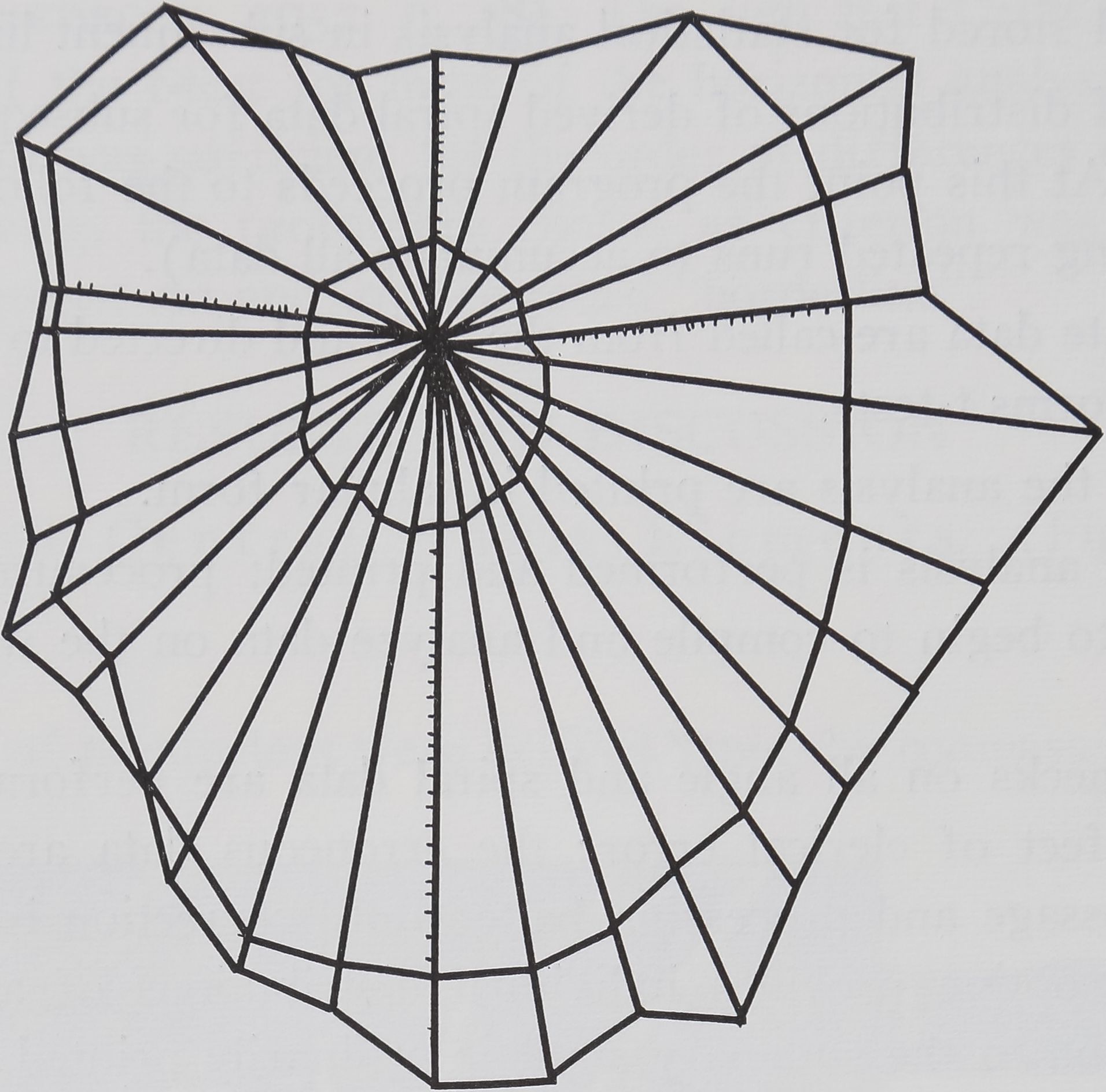
Fig. i. “Control” web of Araneus diadematus Cl. as it is built in the laboratory, sprayed and photographed. Notice the scale in the upper right corner which indicates 2 X 20 mm horizontally and the vertical direction in the original web.
The spider Araneus diadematus Cl. constructs an essentially two-dimensional cartwheel form (Fig. 1). In the finished product of the adult female, about twenty to thirty non-viscid radial threads are evenly disposed about a central hub and are attached to framing threads. A viscid spiral, forming the catching zone, consists of regularly spaced threads occupying an area between, but separated from, hub and frame threads.
In the normal web, the areas not occupied by viscid thread are compara-
100
REED, WITT & JONES
tively small. The hub is placed in a relatively constant position with respect to the perimeter of the spiral area, and a generally oval shape of the spiral zone is maintained, with the vertical dimension longer than the horizontal. While the regularity of the orb has invited speculation about the manner in which it is achieved .(see TinipfciN 1942; Peters, 1953), the disposition of the thread seems td fee vferpfrl^MÉidettt upon the .physical problems involved in construction. At any ¡fát’éy. the ept® rfailt fátowfe in Which there is economy of effort and • >$wkö ‘Wßmm. i blueprints of the web
are programmed spider, Iniaisfenáe. the mass of
the spider determines the thickness jfad~ hence the number
of radiil and ipiral turri^ÉÍÉM>#lfeÍM$íéfe^’ 1$ .BiWMMBfel-ftiilarlv. an in-crease in the production of thread material application of
physogtigjBjlmfe longer thread (Peakaix,
The spider has
built fhd. r^faighf&ty oJro^few:; ¿dái widely
d thread.
‘ in of radii as follows:
fMBHp —‘ 1’h¿g-ir^^HP^t^.aftáMndBphftr clockwise or
… and often one of its second legs can be s^n resting at (the l»b%» feeling
for a space between two threads
evidfntlthis regular tiding and impels tfa$
It seerta^0^E#IÍM^fehte ammaK^^M^K^pr-th^^^^^se of a thread by means of thte. .iirÉK ~Jj|pLvlf-^feE^,d|een: Jifaown that, » radii are destroyed repeatedM during this phase of const ruction, the spider continue to build new radii (Ko Ejb^Mfe ¿L,i® the normal web for the central angles formed b^an^twwMji-hb feffeed tBiMfrof the two adjacent angles (Peters, that elimination of an over
sized angle by pulling neighboring rftdii^^vafd^’ each other prevents the probing spider from putting ^pfpother radips. The measuring function seems to depend upon palpating with the first pair of legs.
Characteristic probing movements d£ the first outer leg have also been noted during the laying of the spiral thread. This phase of building follows completion of all radii and of a non-viscid preliminary spiral. A pause occurs preceding construction of the catching spiral, a different kind of gland provides thread (Peakall, 1964), and henceforward the spider does not modify any of the non-viscid structure.
Jacobi-Kleeman (1953)» ift a detailed photographic analysis of the loco-
MEASURING FUNCTION OF FIRST LEGS OF ARANEUS IOI
motive function of each leg of Arcmeus Cl., observed a groping
of the first leg during spiral placement. The movement was distinguishable from locomotive function. Moreover, the second ipsilateral leg appeared to assume this palpating function in the absence of the first leg.
Hingston, as reported by Saí^rÍH, found the first legs to be
significant both iinfche starting of the preliminar^ spiral and in the spinning of the viscid spiral. Äthedatter ease, the spider appeafird fto place the spiral after locating étee turn ydjtÉMt* firsili lyg-^By remóvmgia portion
of the last fipiKál turn, «Ése* position
of the 1 next f providing rthe first leg
with 4hg tip of Ifee foreleg
was rensiifj.| in/ Pf. made, untidy
web.’?- >’/
It seefeis’. m /<fclgaf> f #j§ ®|p¡®É3S’ obrtrv^&n%othaf, m the ordinary
fafitó#Í5&»{í,spiral, one or and•’pefhap® the
second leg, perform not only a locomotor but also a probing f®áEÉS9i, particularly in the absence of one or
‘bsA in the web pattern.
presence and Compensatory
and adaptive mechanisms the regularity of the web will be
exanÄ^^t’Ä^.ÄSi iÄl^® data were taken
f■ had- been deprived of the first or method of analysis
differs therefore from that of Jacobi-Kleemann (1952). Shrf made a frame ijj^ frame film analysis of web-building y^^Mid6i^t–i”s#ti–£b§ one or 4atiovliii§ and ippe^lst*i^?SaS
Adult- female 4tóiM^^^^p^ps#,jfe.’scaught out
doors. They were;4flf>t dh ih¿ilpféf^«yn#iief•>temperáture and light condítíofi» ¿n.hod%yidthil 20 X 20 X 3.5 inchaluminum frames with glass doors. The ^animals received :WaJter »d¿ily. and housaríliés twice every week. Experience has shown (0Uitt, 19^!, 19631) fhafc ttundfei? these conditions an orb web is built nearly daily during 20-30 minutes at 5:30 A.M.; at this time the light is turned on in the laboratory and the temperature rises steeply.
Every morning the glass doors in the front and back of the frames were removed and the spiders taken out. The webs were then sprayed with Krylon white glossy spray paint. A photograph was taken of each web together with a scale which consisted of three threads 20 mm apart holding a weight (Fig. 1, upper right corner). The scale indicates size and the vertical direction, A black
102
REED, WITT & JONES
box served as background for the illuminated webs. The negatives of the 35 mm high contrast copy film show threads as black lines against a clear background. The films were placed into an enlarger and brought to original size and position with the help of the scale. Measurements were obtained from the projected negatives.
In the first experiments legs were cut off with scissors near the trochanter. It was observed that under those conditions some body fluid leaked out. In order to keep the trauma as light as possible, the autotomy mechanism which exists in spiders was used in later experiments: the animals were carefully held between two fingers and a leg was pulled off with the help of a forceps. Like Jacobi-Kleemann (1953) we did not notice any difference in the consequence of both procedures.
Of the fifteen spiders deprived of one first leg (— 1 spiders), eleven built a sufficient number of webs to warrant statistical analysis; four of the six spiders deprived of the two first ipsilateral legs (—2 spiders) built a sufficient number of webs. The first webs were constructed within the next week, usually the next day, with the exception of one spider who built her first web after 11 days. Altogether 85 control and 102 treatment webs were analyzed.
In order to convert the web into a numerical form representative of the pattern, two forms of measurements were made.
The position of each radius was measured in degrees starting with left horizontal (o° and 360o). The radii do not converge toward a single point but to a locus in the nexus of threads forming the hub; the initial selection of the center is therefore a somewhat arbitrary one. Moreover, the radii, although straight during their construction, subsequently are distorted by the spiral thread. Measurements of radial position therefore were taken at the intersection of the radius with the frame thread.
Having established the compass position of a radius, three points were measured on it: the distance from the center to the first spiral thread, the distance from the center to the outermost spiral thread, and the distance from the center to the nearest frame thread. All measurements were in millimeters. The non-viscid thread forming the hub was not measured.
In addition to these four measures on each radius, the distance of each spiral turn from the center was measured along radii next to the four cardinal compass directions. In parameters derived from the raw measurements, the cardinal compass points are used as representative of a quadrant of the web.
Limitations on accuracy of measurement are imposed by the present techniques. The film is projected to full size. A ruler, pivoted on a compass is centered on the web and readings taken where the projected image intersects
MEASURING FUNCTION OF FIRST LEGS OF ARANEUS IO3
the ruler and compass. Readings are made to the nearest degree or millimeter.
The dimensions of the web, as received by the computer, are shown in Fig. 2.

Fig. 2. A spider web as the computer “sees” it.
Computer Program1).
The first programmed function of the computer, having received the raw measurements of each web, is to generate a number of web parameters. That is, in addition to counting the radii and spiral turns of the web, the computer calculates central angles, areas, total thread length, and certain web proportions for each web (altogether 19 components). When the figures for each web have been obtained, statistical tests are performed on them, comparing any combination of webs selected by the experimenter.
The web analysis program is written in the FORTRAN programming language for an IBM 1620-1311 data processing system. The complete operating program is composed of seven link programs and three subprograms. The functions of their linkages and order of usage are:
1. Initial setting and location of 100 disk records to be employed for the storage of intermediate data from a maximum 40 webs (20 controls and 20 treatment).
2. Reading of data from which central angle parameters are computed;
1) The authors will be glad to provide detailed information on programming on request.
UH
REED, WITT & JONES
intermediate calculations made and stored for analysis by a t-test subprogram.
3. Reading of data from which spiral and associated parameters are calculated and stored for statistical analysis in subsequent links.
4. Forming of distributions of derived spiral data for subsequent Chi-square analysis. (At this point the program proceeds to the following steps only after making repeated runs to accumulate all data).
5. Intermediate data are called from the disk and directed to the subprogram which performs t-tests.
6. Results of the analysis are printed in tabular form.
7. Chi-square analysis is performed and printed; procedure returns to the first step to begin to compile and analyze data on the webs of the next spider.
Sequence checks on all angle and spiral data are performed in order to reduce the effect of clerical errorj the erroneous data are specified in a typewriter message and processing halted until correction is made.
A program option provides individual distributions of angle and spiral data, in addition to the summary statistic ordinarily printed as output.
The program can be. utilized in. two modes: (1) analysis of the webs produced by one spider and (2) analysis of selected webs from many spiders: e.g. the last webs produced before treatment versus the first webs produced after treatment.
Statistical Procedures.
The experimental results will be reported in terms of two types of analysis: A vertical comparison, i.e. comparing each spider’s control webs with its own treatment webs, is of value, in assessing occurrence of effects in an individual. The main computer program was devised with the objective of associating a lesion with a pattern of disturbances in the web structure. The number of webs available in . two conditions places restrictions upon this analysis: the power of the t-test. is limited by the size of the samples (Siegel, 1956).
The data also were analyzed horizontally. The last control webs built by all spiders were compared with the first experimental webs of all spiders and with experimental webs built at least three weeks after delegging. This procedure affords an estimate of general effects which might be obscured by small samples or by the variability of web components in the individual spider. Especially where the effects are subtle rather than extensive alterations of web structure, complementary methods of analysis are useful.
MEASURING FUNCTION OF FIRST LEGS OF ARANEUS
I05
The t-test (Snedecor, 1957, p. 45) is appropriate for comparing the means in both types of analysis, with the exception of the parameters of angle regularity and spiral regularity where the Chi-square technique is appropriate (Snedecor, 1957, p. 18). The sign test (Siegel, 1956) was used in place of the t-test for nidst of the horizontal analyses.; it is a less powerful test but was sufficient for the order of differences observed.
In the following, the probability BH|^^\«riteritin:! was the .01 level, probabilities between .02 and are called “bordérlíii$$H
RESULTS AND i. Radii and;’ Central AijtvJjpl E1 e m e n t s. (Fig. 3)
a. Results.
The first set of parameters dterived ioiäjßotö’s

3 tC *am–C we^ a8 in 1 illustrating the method of measurement of radii.
JDserve the straight lines which do not follow the path of the thread but connect the arbitrary center point with the place where radius and frame thread meet
io6
REED, WITT & JONES
of the radii. Starting from the left horizontal, or West, radius, all the central angles were computed and the radii counted.
Angle regularity: In the normal web, each central angle differs but slightly from its neighbors, although upper (North) sectors tend to have larger angles than do lower sectors. The amount by which adjacent angles differed from each other was taken as an index of regularity of placement of radii.
Table I shows that delegging significantly increased the mean difference between adjacent angles. Two related parameters, number of radii and median angle, were also changed. That the effect is immediate and permanent is shown by testing the last control webs of all spiders against the first webs built after delegging and against webs built at least three weeks later. An additional test, not included in the table, established that early and late treatment webs differed from each other: the number of radii declined significantly.
TABLE i
Radii and central angle elements
| Variable | Control | Experimental | Effect of leg absence on | |||
| X | sd | X | sd | Early webs | Late webs | |
| angle regularity | 4-9° | 1.36 | 7.84o | 2.70 | less regular | less regular |
| number of radii | 2313 | 4.O8 | 18.40 | 309 | fewer radii | fewer radii |
| median angle | 15.30o | 3-13 | 18.83o | 2.72 | larger angle | larger angle |
From each of the 15 spiders one control and one experimental web was chosen randomly and means and standard deviations calculated. To test the onset and permanence of the effects, all last control webs were compared with all webs built immediately after delegging and with webs built at least 3 weeks later. Entries in this case mean that the change was significant at the 0.01 level of confidence.
b. Discussion.
Removal of the first or first plus second leg apparently results in the production of webs with significantly fewer radii, less regularly spaced than in controls. Radii were reduced in number by slightly less than 20% in the average; the median angle was consequently increased significantly. In considering the data spider by spider, i.e. by testing each spider’s total normal production against its total production when minus a leg, four of the -1 spiders failed to show the radial and central-angle effects at the criterion level. While this result may in part be a reflection of the power of the t-test with small numbers, it seems also the case that either variability in web pattern is sufficient to mask effects from the individual spider or that consequences of loss of one of the first legs are not identical for all spiders.
Spiders lacking two ipsilateral legs, however, were all affected. This seems
MEASURING FUNCTION OF FIRST LEGS OF ARANEUS
107

Fig. 4. The distance between sticky spiral turns is measured along radii in the four cardinal directions as indicated in this figure.
to indicate that while the second leg can compensate to some degree for the loss of the first, the absence of both legs makes normal radius construction impossible.
Some reservation is required concerning the use of the t-test in the case of the angle regularity scores. The distribution of differences between adjacent angles in the normal web is skewed, with greater frequencies in the lower difference ranges, zero to five degrees. The t-test requires normality of the underlying measure. As an alternative test, the computer was programmed to prepare separate distributions for all control and all experimental webs for each spider. The hypothesis that the distributions were representatives of the same population was tested by means of the chi-square test. A similar procedure was required in the consideration of spiral regularity, to be discussed below. This test is more sensitive to angle irregularity and verified the findings for the individual spiders.
Results of the comparison between early and late treatment webs indicates
io8
REED, WITT & JONES
that rather than recovering, the ability to build a full number of radii declines further.
2. The Spiral, a. Results.
The number and location of spiral threads were read into the computer for four cardinal directions as shown in Fig. 4. In the normal web, the number of spirals is not uniform in every direction. As can be seen in the means presented in Table 2, a comparatively large number of spiral threads
| TABLE 2 Spiral elements | ||||||
| Variable | Control | Experimental | Effect of leg absence on | |||
| X
Number of spiral turns |
sd | X | sd | Early webs | Late wet | |
| West | 18.13 | 6.40 | 13.67 | 8-93 | no change | no change |
| North | 17.80 | 6.35 | 12.80 | 8.26 | fewer turns | fewer turn |
| East | 23.07 | 7.82 | 16.60 | 7.86 | „ | „ » |
| South | 27.00 | 7.41 | 19.06 | 10.00 | „ V | „ „ |
| Spiral regularity (mm) | 143 | 0.30 | 2.01 | 0.73 | no change | no change |
Means, standard deviations, and late changes. Data were derived and evaluated as in table i.
isr placed in the lower portion of the web, the spider reversing its direction while building additional threads. Webs are built toward one of the upper corners of the square frame; as a measurement convention, the corner at which the ‘web was built was designated West. It should be noted that though the absolute number of spiral turns in each direction decreased after treatment, the ratios between directions stayed the same.
b. Discussion.
Reduction in the number of spirals is slight but significant with the exception of the West quadrant, which is reduced but not to criterion. The effect is observable in the first webs after delegging and is permanent. Absence of a single leg seems sufficient to reduce the number of spirals built.
Results in respect to spiral regularity, i.e. the even spacing of the spiral threads, are less clear. As in the case of the measure for differences between adjacent angles, distances were used to express regularity. The mean dif-
MEASURING FUNCTION OF FIRST LEGS OF ARANEUS IO9
ference for all four quadrants is shown in Table 2. Comparison of the last control webs with the early and late experimental webs does not indicate a reduction in regularity of the spiral. However, the data and tests reported in Table 2 are derived from mean scores. The distribution of differences is narrow and skewed (as in angle differences); hence separate distributions for all control and all experimental webs for each spider were compared by means of the chi-square test. Disturbance at the criterion level was found for all of the -2 spiders and for five of the eleven -1 spiders.
Since the chi-square analysis is performed on all control and all treatment webs of each spider, the temporal course of an effect cannot be gauged. When distributions are prepared for final control webs of all spiders to be compared with final experimental webs, a significant chi-square is not obtained. This finding suggests that spiral regularity may be reinstituted through use of alternative methods of spiral construction. But, for some spiders at least, irregular placement of spirals is a consequence of the absence of the first or the first and second legs.
If the probing function of the first outer leg is of decisive importance in spiral construction, the spider could cqrnpensate for the loss of one front leg by using the intact leg on the outside. This procedure would result in fewer pendulum turns in the spiral, and relatively more circular construction would take place. Such behavior was predicted by Jacobi-Kleemann (1953) but not confirmed by our data. The relative number of spiral turns with respect to direction is unchanged in the treatment webs. The data suggest that the -1 spider as well as the -2 spider rather builds an irregular spiral then changes its procedure of turning back and forth during spiral construction.
3. Areas.
a. Results.
Three areas are calculated, as shown in Fig. 5. The computation of the central area is accomplished by summing the areas of the sectors formed by each central angle and the first spiral thread. Note that the central zone includes the densely-woven hub and the spiral-free zone. The spiral area is computed by subtracting the central zone from the summated areas formed by the central angles and the last spiral turn. A simplifying and not entirely satisfactory assumption is made regarding the frame area. A chord closing the sectors at the distance of the radius-frame intersection is assumed, and the areas of the spiral and center zones subtracted from the summated sectors formed by the central angles and the assumed chord. This procedure yields a frame more regular than the true frame and underestimates the frame area.
I IO
REED, WITT & JONES

Fig. 5. The center area comprises the densely woven hub together with the inner free zone. The catching or spiral area forms a ring around the center area bounded by the innermost and outermost spiral turn. The frame area is smaller in the calculation than in the web because the measuring line connects peripheral ends of radii rather than
following the frame thread.
Borderline statistical significance is reached in the reduction of the three web areas after removal of the forelegs, the effect is significant in all four -2 spiders and for spiral area and center area in one -1 spider. However, all spiders show, in the ratios of the web areas, a relative reduction in the spiral as contrasted with other zones of the web.
b. Discussion.
The spiral area contains the viscid threads and hence constitutes the functional zone of the web. It takes the longest time to build and requires probably a greater amount of body movement than any other part. Its reduction may signify a sparing of effort in the 7 and 6 legged spider. As is evident from the standard deviation in Table 3, the spiral area varies widely in magnitude.
MEASURING FUNCTION OF FIRST LEGS OF ARANEUS
III
TABLE 3 Absolute and relative areas
| Variable | Control | Experimental | Effect of leg absence on | ||
| X | sd | X | sd | Early webs Late webs | |
| frame area (cm2) 148.36 | 43-25 | 132.36 | 72.68 | Borderline Reduction | |
| spiral area (cm2) 327.34 | 115-22 | 216.84 | 108.13 | it tt | |
| center area (cm2) 30.08 | 8.17 | 27-58 | 8.39 | it it | |
| ratio: | |||||
| frame/spiral | 0.491 | 0.185 | 0.500 | 0.227 | Relative Reduction |
| center/spiral | 0.098 | 0.040 | 0.141 | 0.082 | of Spiral |
Means, standard deviations, early and late changes. Data were derived and evaluated as in table I.
4. Position of center and shape of web. a. Results.
Fig. 6 indicates several proportions of the web which were expressed in ratio form. Sable 4 lists the values of the ratios for the random sample and the result! of the statistical tests on the webs.
TABLE 4
Relative position of center and shape of web
| Variable | Control | Experimental | Effect of leg | absence on | ||
| X | sd | – X | sd | Early webs | Late webs | |
| N/S | 0.670 | 0.138 | 0.719 | 0.152 | no change | no change |
| E/W | 0.828 | 0.114 | 0.838 | 0.068 | ”i/, -‘S» tt | tt >t |
| width/length | 0.848 | 0.098 | 0.957 | 0.176 | it tt | tt tt |
Means, standard deviations, early and late changes. Data were, derived and evaluated as in table 1.
In the horizontal measures the larger value was always entered into the denominator in the computer program; the E/W ratio therefore is to be interpreted only as horizontal displacement of the hub.
These findings may be summarized as indicating that the position of the hub with respect to the perimeter of the spiral zone as well as the oval shape stay unchanged.
b. Discussion.
Webs built under the influence of drugs which change central nervous system function (Witt, 1956) can show severe change in the position of
112
REED, WITT & JONES

Fig. 6. The ratios of North to South and East to West radii of the area bordered by the outmost spiral turn provide measures for the position of the hub. North plus South over East plus West diameters is a measure for the oval shape of the catching area.
the center and the shape of the web. The contrast between that finding and the constancy measured after delegging points to a central regulation of web shape and hub position.
5. Thread length and mesh size, a. Results.
The total thread length, viscid and non-viscid, was computed as follows: (1) the lengths of all radii were summed; (2) at the frame distance, an arc closing the sector was assumed and added to the radial sum; (3) the spiral distances measured at the cardinal directions (which were assumed to represent the spirals for four quadrants of the web) were added.
The results of thread length comparisons are shown in Table 5.
MEASURING FUNCTION OF FIRST LEGS OF ARANEUS
113
TABLE s
Total length of thread and average mesh width Variable Control Experimental Effect of leg absence on
| X | sd | X | sd | Early webs | Late webs |
| thread length (m) 13.38 | 5-II | 8.55 | 5.13 | red. length | red. length |
| mesh width (mm2) 66.44 | 12.23 | 80.92 | 26.53 | no change | no change |
Means, standard deviations, early and late changes. Data were derived and evaluated as in table 1.

Fig. 7. The small white quadrangle in the lower part of the web delineates one “mesh”. The average mesh size in a web varies with the relation of thread length to size of the
catching area.
Figure 7 illustrates the area of the web which we have designated mesh size. The functional snare formed by the spiral zone is dependent upon the separation of the threads occupying that area. The number of radii is inversely related to the mean central angle, and was taken as a measure of the average angle. The following formula was employed:
Behaviour XXV
8
REED, WITT & JONES
114
spiral area
(number of radii) X (mean number of spirals)
The formula expresses distribution or density of the threads in the spiral area. In the case of the delegged spider, whether lacking one or two of the forelegs, mesh size, as calculated, remained unaffected.
b. Discussion.
Thread length varies greatly even in the normal web, as is evident in the standard deviation in Table 5. The longest total length in the normal sample, measured by the above procedure was 34.6 meters, the shortest 6.0 meters.
Reduction in total thread length is most clearly evident in the tests performed on the last control versus the early and late experimental webs. The effect is immediate and permanent Every spider’s mean thread length was lower by at least 13% in the experimental condition. There is some sign that the effect is greater for the -2 spiders, but the small numbers do not permit a clear conclusion. The mean reduction in thread length for the -1 spiders was 40% ; for the -2 spiders 58%. If we take thread length also to indicate the length of the way the spider made during web-building, this change would imply the same tendency as the relative decrease in spiral area: decreased total locomotion.
Mesh size is interesting because earlier experimental results (Witt, 1963) indicate that if the spider intends to cover the same area with less thread (less total length available), it resorts to a wider meshed web. The smaller area with unchanged mesh size in our experimental webs again points to a saving of labor rather then to a shortage of thread material.
GENERAL DISCUSSION
The existence of a preformed autotomy mechanism and the frequency with which seven or even six-legged spiders may be found outdoors indicates that the loss of one or two legs does not threaten seriously the survival of the animals. Since the web plays such a prominent role in that survival, it is of interest to examine it for changes in its structure and for evidence of possible compensatory or adaptive behaviors in the delegged spider.
In the matter of pattern, our results confirm that the first or the first and second leg on the same side are essential to a normal arrangement of threads. Every animal except one in our study showed a statistically significant change in at least one of the web parameters which we measured. Loss of these legs — it does not seem to matter whether on the left or right side — results in webs measurably different from those built by intact animals.
MEASURING FUNCTION OF FIRST LEGS OF ARANEUS
II5
Within the limitations set by the size of the samples, it may be inferred that the loss of two ipsilateral legs produces a more pronounced rather than a different effect from that observed in the loss of only the first leg. This finding can be interpreted to mean that the second leg at least partly compensates for the loss of the first leg, with only the absence of the two ipsilateral legs severely interfering with web-building. In contrast to some drug effects (Witt, 1956), the usefulness of the webs as traps for prey was decreased but never eliminated completely.
Some of the pattern changes are attributable to measuring functions served by the legs. Descriptions of web-building (Savory, 1952) seem to indicate that both first legs are necessary for the construction of regularly-spaced radii. Our results lend general support for this conclusion: most -1 spiders and all -2 spiders were unable to establish the width of the central angles with the precision observed in their control webs. The spiral tended to become irregular also, confirming the assumption of Jacobi-Kleemann (1952) that the first leg, or in its absence the second, has a probing function to perform during spiral construction. However, from time to time after delegging, a single web with a regular spiral was observed, ‘suggesting that an alternative mechanism could substitute for the lost function. Regular spiral spacing can be achieved Snply by making the shortest bridge from radius to radius (Witt, 1954). Such a mechanism may become predominant in the absence of a leg. In any event, regulation of spiral placement ‘seems more complexly determined than does central angle regularity.
Other changes in web pattern may be the consequence, not simply of loss of measuring ability, but of severe handicap in locomotion. While Jacobi-Kleemann observed unaltered gait or adequate compensation of gait in delegged spiders, some changes in the web may be interpreted as alteration in the spider’s motor behavior. The decrease in size of the whole web, the relative decrease in the size of the spiral area (which is most complex and difficult to construct), and the preservation of mesh size, all point toward an economy of effort. This pattern change is in contrast to that observed when material available for thread production was diminished (Witt, 1963) : under that circumstance, the animals built full-sized webs with wider meshes. Interpretation in terms of . limited labor also applied to the reduction in number of radii: lacking the measuring function alone, it is not clear that the spider necessarily would produce fewer radii.
If the first web is viewed as adaptation to the new condition (6 or 7 leg-gedness), our experiments show that no further adaptation occurs. The appearance of regular spirals noted above was an occasional phenomenon rather than one of a series indicative of recovery. In no instance did a web
Fig. 8. Upon loss of at least one of the forelegs, there is a general decrease in the size of the web of Araneus diadematus Cl. The total length of thread ‘used to build the web is less than it is in control webs. The web may become smaller in total area, or the thread may be distributed in fewer radii and spiral turns or both. The radii are less regularly spaced than normally, the spirals may or may not be irregular in placement. More of the available space of the web is left, open, i.e. not filled by the viscid spiral. The oval shape of the sticky spiral area remains unchanged, as does the relative position of the hub within it.
a. Control web.
b. Web of the same spider after it had been severely disturbed by elimination of a leg.
c. Control web.
d. Web built by the same spider after it had been less severely disturbed than the spider in (a) and (b) by the same operation. Observe that in comparing (b) and (d) it can be noticed that while qualitatively the changes are alike, not all spiders showed the same quantitative disturbance.




MEASURING FUNCTION OF FIRST LEGS OF ARANEUS
117
parameter show a more severe disturbance directly after the operation than it did later. In one component — the number of radii — the reduction became greater with time.
Because animals differed in the number of webs available for statistical comparison, it is not possible to draw extensive conclusions from the appearance of statistically different changes in one animal and not in another. However, evidence was obtained of differences between individual spiders in their reaction to the operation (Fig. 8). Two animals missing only one leg showed continuous severe changes comparable to those of animals missing two legs. One spider (which built nine webs after being deprived of one leg) did not show any statistically significant alterations. These differences in reaction had no apparent correlation with variables such as body weight or age. The variability observed in the spiders suggests that it may be advisable to impose homogeneity on the sample used in investigating experimental effects, i.e., by selecting animals which build controls webs with parameters within normal ranges.
The detailed and extensive examination of web pattern which is made feasible by the computer enlarges the study of the behavior of the spider. Means are provided for examining hypotheses on an adequate sample of webs. For example, our data did not confirm the generality of the observation made by Jacobi-Kleemann (1953) on periodic spiral changes in the web of a delegged spider. The phenomena which she observed in that single web appear to be not causally related to the absence of the leg.
The employment of the computer has a practical consequence in the use of the web as a pharmacological test. The evaluation of drug effects through the measureable changes certain drugs produce in the web pattern has suffered from the amount of labor involved in the analysis of web geometry. By means of the computer, a great number of data points (about 600 per web) derived from several webs built by a spider (up to 20 control and 20 treatment webs) can be processed and evaluated statistically in a relatively brief time. Detection of subtle changes depends especially upon the number of figures available. It is now possible to contemplate the orb web not only as an interesting structure but as a vehicle for the experimental study of physiological and behavioral phenomena.
SUMMARY
Spiders deprived of one first leg or two front legs on the same side built webs changed in specific ways and never recovered.
Web components were changed from control values to a statistically significant degree as follows: angle regularity and the number of radii decreased, the median central angle increased in size; the number of spiral turns and, in some cases, the spiral regularity decreased; the spiral area decreased relative to frame and central areas; the thread
REED, WITT & JONES
Il8
length decreased. No change could be detected in web shape or in the position of the hub. It is concluded that the first leg has a measuring as well as a locomotor function, that the second leg can partly substitute for that measuring function. Web changes following delegging reflect loss of normal adequacy in both of these functions. No evidence of right-left differences in leg function were found. All adaptation to the absence of one or two legs had occurred in the first web after the operation.
The variation in control webs and the differential severity in reaction to delegging require evaluation of several animals and many webs; such extensive evaluation is made feasible by computer analysis of an array of points defining i web.
REFERENCES
Christiansen, A., Baum, R. & Witt, P. N. (1962). Changes in spider webs brought about by mescaline, psilocybin and an increase in body weight. — J. Pharmacol., 13b, p. 31-37.
Jacobi-Kleemann, J. (1953). Über die Lokomotion der Kreuzspinne Ar anea diadema beim Netzbau (nach Filmanalysen). — Zschr. vergl. Physiol., 34, p. 606-654. Koenig, M. (1951). Beiträge zur Kenntnis des Netzbaus orbiteier Spinnen.— Zschr.
Tierpsychol., 8, p. 462-492.
Peakall, D. B. (1964). Composition, function and glandular original/the silk fibroins of the spider Araneus diadematus Cl. — J. exp. Zool. 156, p. 345-352.
Peters, H. M. (1951). Untersuchungen üffir die Proportionierung im Spinnen-Netz.
— Zschr. Naturforschg. 6b, p. 90-107.
—— (1953). Weitere Unt^puchungen über den strukturellen Aufbau des Radnetzes der Spinnen. — Zschr. Natur forschg. 8b, p. 355-370.
Savory, T. H. (ig52).g|rhe spider’s web. — Frederick Warne & Co., London.
Siegel, S. (1956). Nonparametric statil|l!|l for the behavioral menees. ^HMcGraw Hill, New York, p. 68.
Snedecor, G. W. (1957).. Statistical methods. — The Iowa State College Press, Ames, Iowa.
Tilquin, A. (1942). La téjle géométrique des araignées. — Presses Universitaires de France, Paris.
Witt, P. N. (1952). Ein einfaches Prinzip zur Deutung einiger Proportionen itn Spinnennetz.^® Behaviour 4, p. 172-189.
– (1956). Die Wirkung von Substanzen auf den Netzbau der Spinne als biologischer
Test. — Springer Verlag, fieidelberg.
– (1958). The identifieftion of small quantities of hallucinatory substances in body
fluids with the spider test.||¡|¡ In PBchopathMogy, a source book, ed. C. F. Reed, Harvard University Press, Boston.
– (1963a). Environment in relation to behavior of spiders. — AMA Arch. Envir.
Health, 7, p. 10-18.
– (1963b). Effects of physostigmine on spiders’ web building behavior and thread
production. — The Pharmacologist, abstract.
– & Baum, R. (i960). Changes in orb webs of spiders during growth. — Behaviour
16, p. 309-318-
-, Reed, C. F. & Tittel, F. K. (1964). Laser lesions and spider web construction.
— Nature 201, p. 150-152.
ZUSAMMENFASSUNG
Spinnen, denen ein erstes Bein oder beide Vorderbeine einer Seite entfernt worden waren, bauten Zeit ihres Lebens spezifisch veränderte Netze.
Die folgende Masse waren statistisch sicher verändert: die Zahl der Radien war vermindert, der Mittelwert der Differenzen zwischen benachbarten Winkeln war ver-
MEASURING FUNCTION OF FIRST LEGS OF ARANEUS II9
grössert, die Umgänge der Klebspirale waren weniger zahlreich und hatten ungleich-massige Abstände. Die Gesamtlänge des Fadens war verkürzt. Aber die Lage der Nabe und die Netzform waren unverändert.
Aus diesen Befunden ist zu schliessen, dass das vorderste Bein ausser zur Ortbewegung auch zum Messen dient und hierin teilweise durch das zweite Bein ersetzt werden kann. Die beschriebenen Veränderungen des Netzes nach Entfernung eines oder beider Vorderbeine einer Seite spiegeln Veränderungen in beiden Funktionen wider. Dagegen waren keine Unterschiede zwischen Netzen von Spinnen ohne rechtes bzw. ohne linkes Vorderbein gefunden. Soweit überhaupt eine Anpassung an den neuen Zustand zu beobachten war, hatte sie sich im ersten nach der Operation angelegten Netz schon voll ausgewirkt. Die Streuung bei den Kontrollnetzen und Unterschiede in dem Störungsgrad durch Beinverluste zwingen dazu, viele Netze mehrerer Tiere für einen solchen Versuch auszuwerten. Für so ausgedehnte Analysen der wesentlichen Punkte im Netzmuster war der Computer unentbehrlich.