Images Collection




View this article in Search Friendly Plain Text
NOTE: This plain text article interpretation has been digitally created by OCR software to estimate the article text, to help both users and search engines find relevant article content. To read the actual article text, view or download the PDF above.
nim. Behav., 1970,18,478-484
ORB WEB CONSTRUCTION: INTERACTION OF SPIDER (ARANEUS DIADEMATUS CL.) AND THREAD CONFIGURATION
By PAUL J. PETERS*
North Carolina Department of Mental Health,Research Division, and North Carolina State University, Department of Zoology, Raleigh, North Carolina
rb web construction by the ^cross-sgider’ produce a functional web. The degree of latitude
ANIMAL
BEHAVIOUR
LONDON
BAILLIÈRE, TINDALL & CASSELL
Reprint No. 228 from Animal Behaviour, 18, 3, August, 1970
Anim. Behav., 1970,18,478-484
ORB WEB CONSTRUCTION: INTERACTION OF SPIDER CARANEUS DIADEMATUS CL.) AND THREAD CONFIGURATION
By PAUL J. PETERS*
North Carolina Department of Mental Healthy Research Division9 and North Carolina State University y Department of Zoology, Raleigh, North Carolina
Orb web construction by the ‘cross-spider’ Araneus diadematus Clerck goes through three arbitrary but distinct phases: the frame and radii (Phase I), the auxiliary spiral (Phase II), and the viscid or catching spiral (Phase III) (Fig. 1). Details of web construction have been adequately described elsewhere (Tilquin 1942; Savory 1952; Witt, Reed & Peakall 1968). One kind of silk is utilized in Phases I and II, and in Phase III the baseline silk receives a coat of viscid substance (Peakall 1964).
To build such a complex structure, a be-haviourally stereotyped animal must follow a well-defined programme (Carthy 1965, p. 10). Spiders raised in isolation so that they could not construct webs, built adult webs the first time upon release, showing that a programme for the web pattern is established without previous web building experience (Petrusewiczowa 1938; Mayer 1953; Szlep 1961).
Studies such as Eberhard’s (1969), where an attempt was made to plot a web by computer, imply that stimuli from the web trigger subsequent behaviour. Others, in contrast, had found that a spider will not replace an auxiliary spiral if it was destroyed during or just after its placement (Hingston 1920; Koenig 1951). This failure to fill-in a missing structure means that the spider executes some fend of pre-programme independent of external cues. The temporal order, at least for the phase sequence, cannot be altered.
A third type of experiment indicates that a spider does respond to external cues up to a point, but then ignores those cues and goes to the next phase. A radius burned out just as it was placed was replaced by the spider about twenty-five times (the total radius number is usually twenty-five to thirty-five). However, at this point the missing radius was ignored and the spider went on, leaving Phase I incomplete (Hingston 1920; Koenig 1951; Reed 1969).
It was suggested above that a spider must execute an already existing programme to
•Present address: Zoology Department, University of Texas, Austin, Texas 78712.
produce a functional web. The degree of latitude for the spider in following this programme, and indeed the cues used by the spider for execution of it, are not clear. Natural selection will have insured that the usual programme results in a good web, but the spider is so dependent on its web that something less than the best would be better than nothing at all.

Fig. 1. Partially completed web with major structural components labelled. The frame and radii, as well as the hub (Phase I), are constructed together. The auxiliary spiral (Phase II) is built from the hub outward, and then, after a pause and change of direction, the viscid spiral (Phase III) is built from the periphery to the hub. The spider has removed part of the auxiliary spiral as it placed the first seven turns of viscid spiral.
The present paper attempts to establish the relative importance of external and internal cues used by A. diadematus in web building. The rigidity of the web-building programme in regard to the phase sequences is also investigated. The experiments described are the
478
479
PETERS: ORB WEB CONSTRUCTION
destruction of a phase just as it is completed, and transferring a spider during web building to the partial web of another spider.
Methods
Subjects
Spiderlings of A. diadematus, obtained from New York (Mr Leonard Pankhurst, 204 Stroud Street, Canastata, New York), were raised in a wire-mesh box containing a large number of gnats (Hippelates pusio). When the spiders began to build webs, they were individually separated into small jars; a supply of Drosophila and water was maintained. As soon as they could catch a housefly (Musca domestica) the spiders were housed in aluminium cages (Breed et al. 1964) and fed two flies per week.
Normally, spiders build webs in the temperature minimum just before sunrise (Spronkl935). The onset of building appears to be influenced by light and temperature (Witt 1963) so these factors were used in an attempt to delay web building. Automatic temperature and light control provided a 16 hr day and 8 hr night which were delayed about 3 hr from the normal sunrise and sunset. A room humidifier was in operation at all times. An apparent shift in time of building was not realized with this procedure, but an asynchronous delay was achieved by placing the spiders in a refrigerator for 2 hr at 5°C.
Spiders were first used in experiments 3 to 4 months after emergence from the egg sack. One could expect these spiders to moult two or three times before becoming non-moulting adults. A few adults were also used as subjects but they are not as easy to manipulate as the younger ones. Weights ranged from 18 to 433 mg (mean = 83 mg, median = 54 mg), and the lengths of the first leg were 7 to 20 mm (mean = 12*6 mm, median = 12-0 mg). Sexual dimorphism is marked, the male being smaller and lighter in weight than the female. Only females were observed since the adult male tends to be a wanderer and probably does not build webs.
Experiments
Spiders were kept continuously in the closed aluminium frames. For an experiment, twelve frames were opened and suspended overnight by strings and wire hooks (Fig. 2). Any existing web structures were removed. Glass tubing smeared with petroleum jelly prevented spiders from escaping upward. Dropping downward was common, but the spiders usually stopped a few

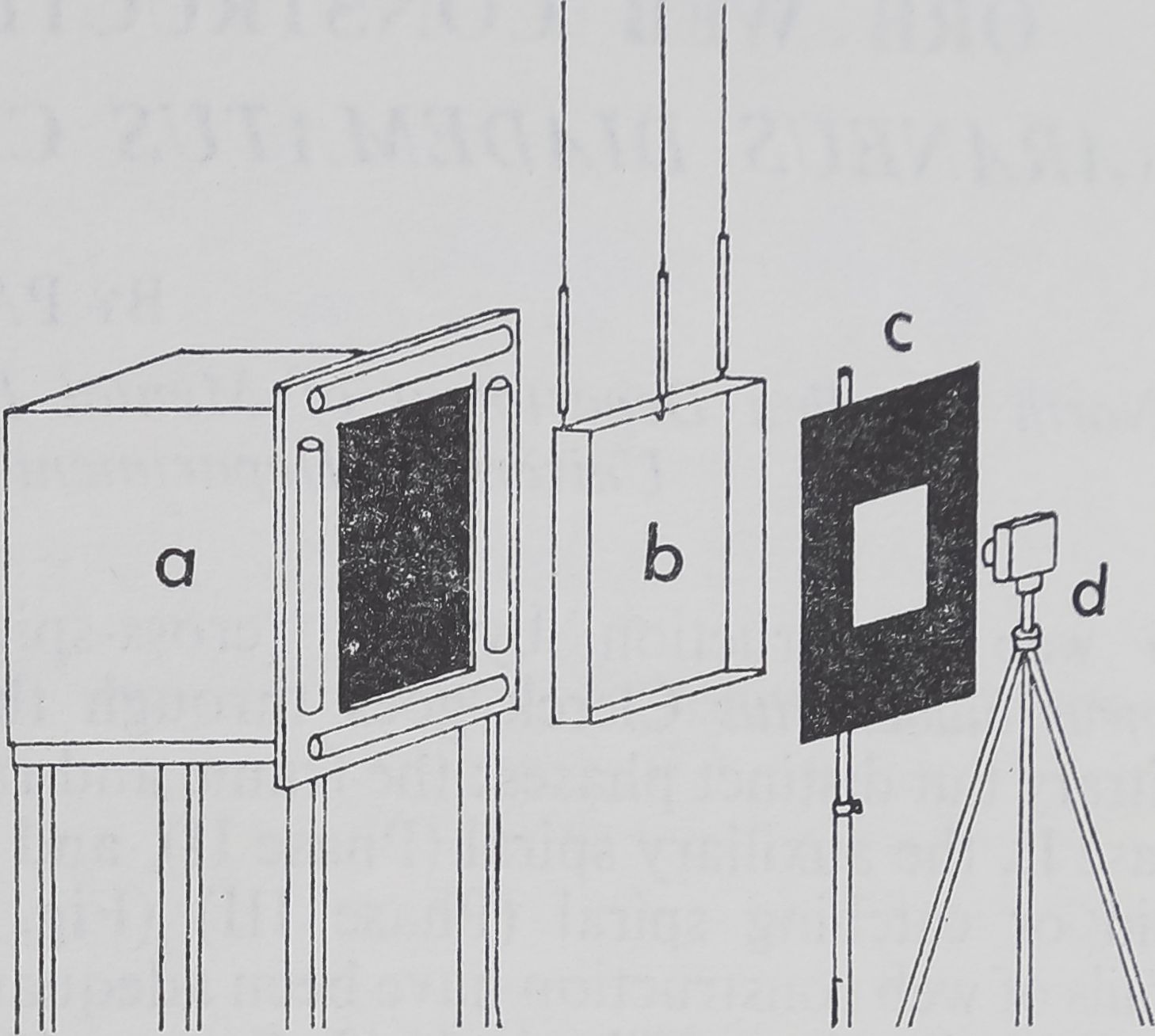
Fig. 2. Diagram showing the relationship of the background and lights (a), aluminium cage frame (b), light-mask (c) and camera (d). The actual distance between any two components can vary to provide the best web illumination. Drawing is not to scale.
centimetres from the floor and then went back to the aluminium frame. Some escaped by wafting out lateral bridge threads.
The following morning, lights in this room were turned on 1 hr earlier than usual. Surveillance of the spiders took place through a window in a hallway, as spiders before building are exceptionally sensitive to disturbances. After web building had commenced, the room was carefully entered and the black-box background plus light-frame (Fig. 2) were moved behind the building spider. The spider could be removed by forcing it to run into a plastic vial, which was then capped. The spider was retained for about 15 min.
Manipulation of the spider or web took place during the time the spider was removed. When appropriate, threads in the incomplete web were removed by burning through with a battery-powered surgical cauter. The bisected threads contracted leaving only a short, loose end at the radius.
Photographic Technique
To a black-box background such as that described by Witt (1956), four flourescent tubes (20 W each) were added (Fig. 2). The legs of the box were mounted on large rollers. A mask placed in front of the subject admitted to the camera only light reflected by threads. Being mobile, the entire set-up could be moved without greatly disturbing the animals. This technique is similar to that of Langer & Eberhard (1969), and can readily be adapted for cine photography.
Photographs were made with 35 mm Kodak Panatomic X film (ASA 32). A measuring device
PLATE X

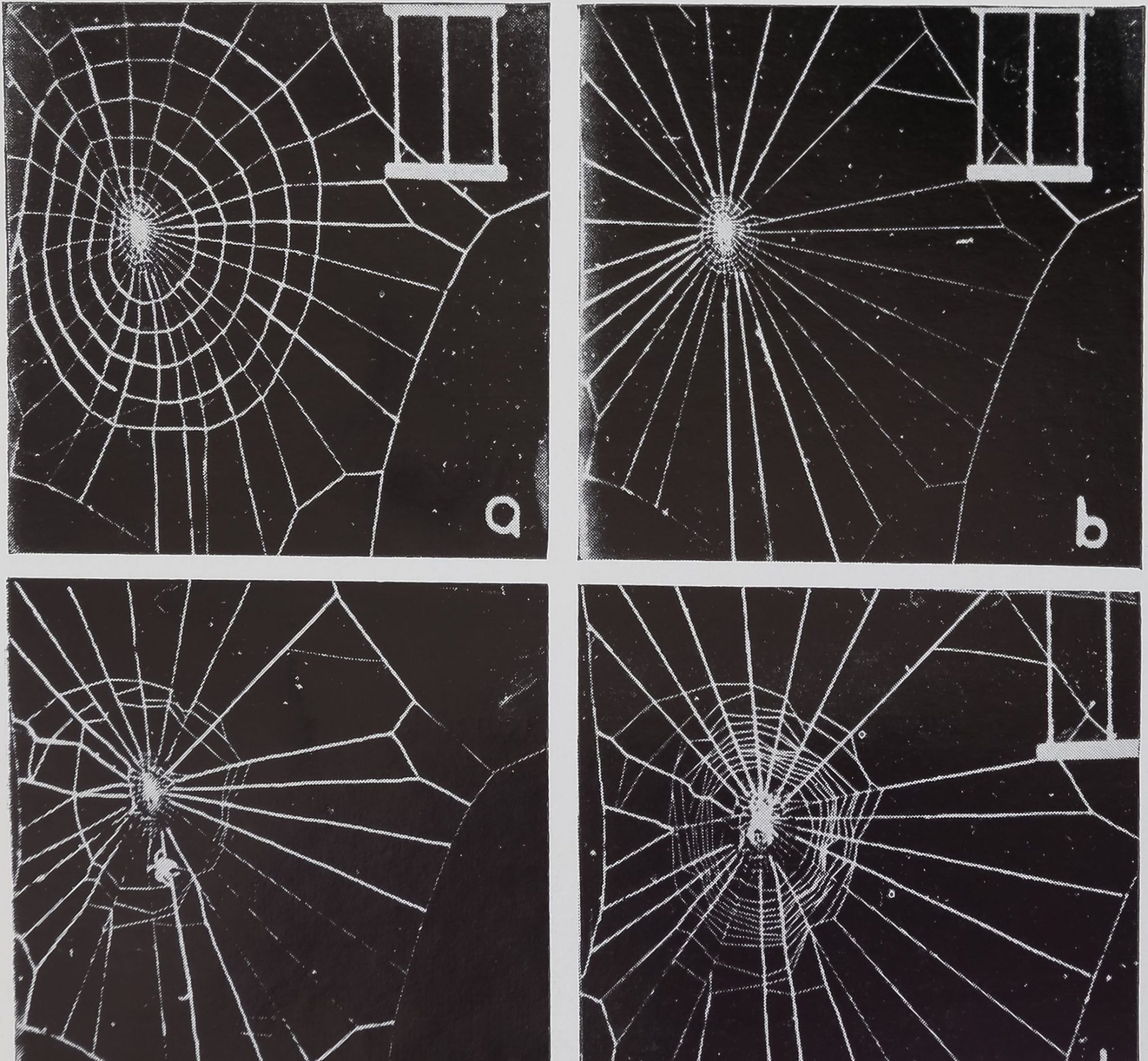
Fig. 3. Stages in a web reversion experiment in which the spider built a second but normal sized auxiliary spiral. The spider was taken off after completing (a). The cut ends of the first auxiliary spiral can be seen in (b). The returned spider builds a new auxiliary spiral (c), which can be directly compared with the first in (a). It then finished a regular catching spiral in (d).

Fig. 4. Stages in web reversion experiment in which the spider built the viscid spiral without repeating the auxiliary spiral. The spider was taken off when the web was at stage (a), the web was changed to (b), and the returned spider continues as in (c). The irregular result in (d) is in striking contrast to the usual finished web (Plate X, Fig. 3d).
c
Peters, Anim. Behav.t 18, 3
PLATE XI

Fig. 5. Stages in a web reversion experiment in which the same spider built a second but smaller auxiliary spiral. Compare the size of the spiral in (c) with that in (a). The finished viscid spiral in (d) does not fill the web frame to as great an extent as does the control animal.

Fig. 6. Catching area built by a smaller spider placed on the completed auxiliary spiral of a larger spider, the arrow indicating the first viscid threads placed by the larger spider before its removal.
Peters, Anim. Behav18, 3
ANIMAL BEHAVIOUR, 18, 3
480
was constructed of three threads, 20 mm apart, and a metal bar. This allowed for scaled reproduction from the negative, and the bar indicated the horizontal plane. Magnets were used to attach the device to the cage frames.
Results
Manipulation of the spider and threads during web building did not significantly decrease the frequency of web completion when compared to controls (Table I). The control group was removed at the end of Phase II and then replaced on their unaltered webs after 15 min. The building frequencies of the three groups were compared by x2 test for K independent samples (Siegel 1956, p. 104).
Interruption did have an effect since 50 per cent of the controls did not finish the web. Had they not been interrupted, which included removal from the web and a delay, one would expect 100 per cent completions. The significant finding here, is that reverting to an earlier phase, and transferring spiders during building caused no greater decrease in web completion than interruption alone.
Web Phase Reversion
Web reversion experiments consisted of removing the spider at the end of Phase II (Plate X, Fig. 3a), burning the auxiliary spiral through (Plate X, Fig. 3b), and replacing the spider on the reverted web (Plate X, Fig. 3c). In seven out of eleven trials (Table I) the spider completed the web after interruption and removal of the auxiliary spiral. Four spiders did not accept the altered web; their building activities stopped for the rest of the day. Of the seven that completed a web, five repeated the auxiliary
spiral and two built the viscid spiral without repeating. The differences between the finished webs of those spiders that repeated (Plate X, Fig. 3d) and those that did not (Plate X, Fig. 4d) is striking. One photograph (Plate X, Fig. 4c), shows a spider in construction of a viscid spiral while the auxiliary spiral is missing.
The repeated auxiliary spiral was not always the same size as the first, as suggested by the reduced size of the viscid spiral (Plate XI, Fig. 5d). A direct comparison of the first (Plate XI, Fig. 5a) and second (Plate XI, Fig. 5c) auxiliary spirals reveals that the second spiral was smaller.
Some spiders did reproduce the first spiral. A viscid spiral that filled the frame would indicate that a full-sized auxiliary spiral had been repeated. A direct comparison of the first spiral (Plate X, Fig. 3a) with the second (Plate X, Fig. 3c) shows an almost identical repetition of the auxiliary spiral.
Web Transfers
Transfer experiments involved removing a spider at the completion of Phase I or Phase II, and then placing it on another partial web in the same phase. Only one spider failed to resume web construction. The remaining four negative completions (Table I) were negative only in the sense of the criterion imposed for the frequency-of-completion test; viz. completion of a web on the existing structure. These four did not meet the criterion, but they did build a complete web by starting again.
Quantitative evidence for size of frame-areas was obtained by measuring all radii from hub centre to frame and calculating the median for both the original and foreign webs (Table II).
Table I. Frequency of Web Completion by Spiders that had a Web Phase Reverted or that had been Transferred to Another Partial Web is Compared with a Group that had been Interrupted
Only (Control)
| (+> | (-> | Total obs. | |||
| Obs. | Exp. | Obs. | Exp. | ||
| Control | 6 | 6-86 | 6 | 514 | 12 |
| Reverted | 7 | 6*29 | 4 | 4-71 | 11 |
| Transferred | 7 | 6*85 | 5 | 5*14 | 12 |
| Total obs. | 20 | 15 | 35 |
X2 = o-446, Prob. H0 = 0-80.
Expected values are based on the marginal totals. Plus (+) indicates web completion on the existing threads; those that started again from the beginning or ceased all activity were scored as minus (—).
481
PETERS: ORB WEB CONSTRUCTION
Table IL Median Radius Length in the Frame Area of the Spider’s Own Partial Web, Foreign Partial Web, and in the
Catching Area Built on the Foreign Partial Web
| Spider’s
weight (mg) |
Own | Foreign | Catch | |||
| Frame
(mm) |
No. of radii | Frame
(mm) |
No. of radii | (mm) | ||
| Phase I transfers | 37 | 91 | 20 | 102 | 24 | 65 |
| 63 | 102 | 24 | 91 | 20 | s.a. | |
| 28 | 61 | 26 | 100 | 27 | 71 | |
| 36 | 100 | 27 | 61 | 26 | s.a. | |
| 32 | 95 | 23 | 144 | 24 | s.a. | |
| 72 | 144 | 24 | 95 | 23 | s.a. | |
| Phase II transfers | 185 | 103 | 19 | 142 | 20 | 98 |
| 226 | 142 | 20 | 103 | 19 | 86 | |
| 23 | 53 | 23 | 72 | 24 | 52 | |
| 37 | 72 | 24 | 53 | 23 | 46 | |
| 134 | 191 | 30 | 184 | 27 | — | |
| 165 | 184 | 27 | 191 | 30 | 158 |
The number of radii in the original and foreign webs are shown, s.a. = started again.
The median of the radial lengths from hub to last viscid spiral turn of the finished web gives an indication of the size of the catching area (hub + free zone + viscid spiral) as it was built on the foreign, partial web.
To determine whether the catching area was large, small, or normal in size, the per cent of the foreign and original frame areas filled by catching area was calculated. Using median radius lengths (Table II), the area encompassed by the projected circle was calculated. The size of the catching area divided by the area of the foreign and original frames yields the fraction filled by the catching area. Table III shows per cent of area filled.
Data from webs built overnight without interruption were used to establish the normal size of the catching area. In twenty webs, containing nineteen to thirty radii (mean = twenty-four), the mean catching area filled 73’1 per cent ± sd 6-7 per cent of the mean framed-in area.
Every spider which started over again (Table III) located the new web in that corner of the cage in which the previous web had been. In some cases the existing frame threads were partially
incorporated into the new web. The single Phase I spider transferred to a larger partial web, that started again, cut an area out of the large web and built there. The fact that this spider weighed less than one-half as much as the original builder may explain such behaviour.
If a spider accepted the auxiliary spiral of a web larger than its own, it still did not put in a normal sized catching area. For example, one heavier spider had placed three turns of viscid thread before it was removed (Plate XI, Fig. 6). The lighter spider placed on this partial web used the same auxiliary spiral, but its outer thread did not reach as far toward the frame as the heavier spider’s. The lighter spider probaby had shorter legs, and a mechanical limitation was in effect.
One transfer resulted in a normal sized catching area for the foreign and original frames. This could be expected from the fact that the two frames, only 7-0 mm different in median radial length (Table II), were about the same size.
Discussion
Any conclusions from these experiments are
ANIMAL BEHAVIOUR, 1 8, 3 482
Table m. Size of the Catching Area Built by a Spider (B) Transferred to the Partial Web of Another Spider (A)
| Spider A:
weight of initial builder (mg) |
Spider B:
weight of spider that completed web (mg) |
A/B | Phase, and foreign web size compared to own | Per cent foreign web filled | Relative
size |
Per cent own web would be filled | Relative
size |
| 37 | 63 | 0-59 | I-smaller | s.a. | — | — | — |
| 28 | 36 | 0-78 | I-smaller | s.a. | — | — | — |
| 32 | 72 | 0-44 | I-smaller | s.a. | — | — | — |
| 72 | 32 | 2-25 | I-larger | s.a. | — | — | — |
| 63 | 37 | 1-70 | I-larger | 40-6 | Small | 51 -0 | Small |
| 36 | 28 | 1*29 | I-larger | 50-4 | Small | 135*5 | Large |
| 226 | 185 | T22 | II-larger | 47-6 | Small | 90*5 | Large |
| 37 | 23 | 1*61 | II-larger | 52-2 | Small | 96*3 | Large |
| 134 | 165 | 0-81 | II-same | 68-4 | Normal | 73*7 | Normal |
| 185 | 226 | 0-82 | II-smaller | 69-7 | Normal | 36*7 | Small |
| 23 | 37 | 0-62 | II-smaller | 75-3 | Normal | 40*8 | Small |
Both webs were at the phase indicated. The catching area is compared to the expected size for both partial webs. Normal catching area of a reference group (iV=20) filled 73*1 ± sd 6*7 per cent of the framed-in area. Note that spider weight corresponds well with web size.
limited by the small number of individuals in each experiment. There is no a priori reason to expect behavioural differences due to age, as the webs of ‘juvenile’ spiders are hardly distinguishable from those of adults in regularity or amount of detail (Witt & Baum 1960). My transfer experiment (putting an animal on another animal’s web) cannot be directly compared to those of Petrusewiczowa (1938). Her spider transfers were of four types: (i) to more advanced phases, (ii) from no web to one partially completed, (iii) to less advanced stages, and (iv) from finished to unfinished webs. The transfers described in the present paper were always to another partial web at the same stage of completion.
The spiders that completed their webs after the auxiliary spiral had been destroyed either repeated the structure or continued without repetition. Some of the second spirals were smaller than the first, and some were full-sized. Comparable results were obtained by Hingston (1920), Koenig (1951) and Reed (1969). They found that a spider will replace a missing radius about twenty-five times and then leave an incomplete radius phase to finish the web.
Such results suggest that something is pro
gressively depleted during web construction. Silk is an obvious candidate for the substance depleted so that a monitor on the amount of silk left could provide feedback information to the central nervous system. However, such an idea is speculation at this time.
The high degree of irregularity in those webs which were completed without an auxiliary spiral (Plate X, Fig. 4d) points to the important role which this structure plays in catching-spiral construction. The auxiliary spiral acts as a temporary stabilizer for free radii, preventing their being severely displaced by the weight of the animal ; it also serves as a bridge, particularly in the periphery, for the spiral-laying spider. In both functions it aids in the regular spacing of the catching thread.
Transferring a spider to another’s partial web shows that the size of the catching area is determined by the size of existing structures (radius length, central angles, and so on) within limits of the building spider’s size. If size were determined strictly by existing structures, a transferred spider would build a catching area normal in size for the foreign structure. This was true only for exchanges to smaller webs where the auxiliary spiral was already present. Here, the frame
483
PETERS: ORB WEB CONSTRUCTION
threads of the smaller web limited the larger spider within the given area, thus producing a normal sized catching area. That spiders will build within an imposed thread boundary was illustrated by Koenig (1951).
A large, foreign web, with one exception, resulted in a larger than normal catching area for the spider, but smaller than normal for the existing structure. The influence of the spider, in this case, was shown probably to be a mechanical one due to locomotor difficulties in the periphery of the large web.
All heavier spiders placed on the radii stage built by lighter spiders destroyed the existing structures and rebuilt from the beginning. An obvious explanation is that the radii and frame threads alone were too weak, thin or unstable to support the heavier spider’s weight. Such an assumption is supported by the observation that heavier spiders generally produce thicker threads (Witt & Baum 1960J- Christiansen, Baum & Witt 1962). One spider which weighed less than one-half of the first builder started again when given the larger web; it probably could not manoeuvre on the much larger structure.
It seems reasonable to conclude that the innate, neural programme for web construction by A. diadematus is expressed as responses to external stimuli, viz. previously laid threads. There may also be higher order stimuli arising from internal conditions that modify the normal responses to external stimuli.
Summary
The functional significance of stimuli from the spider as opposed to those from the web was tested. The approach was to alter the normal web building sequence, and to transfer a spider during building to another partial web at the same stage of completion.
Web construction proceeds through a sequence of three phases. It was found that a spider can repeat a phase, thus taking cues from the web, or it can go on, independent of fit to the structure. Phase repetition indicates that the phase sequence is not centrally executed without regard to external stimuli. A repeated auxiliary spiral was either equal in size to the first, or smaller; the latter resulted in a correspondingly smaller catching area (hub + free zone + viscid spiral). In regard to the different sizes, or nonrepetition of the auxiliary spiral, it was suggested that some substance (silk?) was being depleted during the course of construction.
With one exception, a transferred spider built a catching area corresponding to the size of the structure on which it was built. Larger spiders built within a small foreign frame if an auxiliary spiral was present; they started completely again if the spiral was lacking. Smaller spiders on large foreign webs built larger than normal catching areas but did not fill the frame area.
It was concluded that external stimuli act as triggers for subsequent behaviour. Also, it is possible that stimuli from the milieu intérieur of the spider modify the normal responses to thread configuration.
Acknowledgments The author wishes to acknowledge the guidance and encouragement of Dr Peter N. Witt throughout this study. Gratitude is extended to Dr Witt and Dr David E. Davis for reading the manuscript. This work was supported in part by National Science Foundation Grant No. GB-6246X to P. N. Witt.
REFERENCES Breed, A. L., Levine, V. D., Peakall, D. B. & Witt, P. N. (1964). The fate of the intact orb web of the spider Araneus diadematus. Cl. Behaviour, 23,43-60. Carthy, J. D. (1965). The Behaviour of Arthropods. San-Francisco: W. H. Freeman.
Christiansen, A., Baum, R. & Witt, P. N. (1962). Changes in spider webs brought about by mescaline, psilocybin and an increase in body weight. J. Pharmac., 136, 31-37.
Eberhard, W. (1969). Computer simulation of orb web construction. Am. Zool., 9(1), 229-238. Hingston, R. W. G. (1920). A Naturalist in Himalaya.
London; H. F. & G. Willoby.
Koenig, V. M. (1951). Beitrâge zur Kenntnis des Netzbaus orbiteler Spinnen. Z. Tierpsychol., 8, 462-493. Langer, R. M. & Eberhard, W. (1969). Laboratory photography of spider silk. Am. Zool., 9(1), 97-102,
Mayer, G. (1953). Untersuchungen Uber Herstellung und Struktur des Radnetzes von Aranea diademata und Zilla x-notata mit besonderer BerUcksich-tigung des Unterschiedes von Jugend und Alter-snetzen. Z. Tierpsychol., 9, 337-362.
Peakall, D, B. (1964). Composition, function and glandular origin of the silk fibroins of the spider Araneus diadematus Cl. J. exp. Zool., 156, 345-352.
Petrusewiczowa, E. (1938). Boebachtungen Uber den Bau des Netzes der Kreuzspinne (Aranea dia-dema L.). Trav. Soc. Sci. Lettr. Wilno (Trav, Inst. Biol. Wilno No. 9), 13, 1-24.
Reed, C. F. (1969). Cues in the web-building process.
Am. Zool., 9(1), 211-221.
Savory, T. H. (1952). The Spider’s Web. London: Frederick Wame.
Siegel, A. (1956). Nonparametric Statistics for the Behavioural Sciences. New York: McGraw-Hill. Spronk, F. (1935). Die Abhàngigkeit der Netzbauzeiten der Radnetzspinne Epeira diademata und Zilla x-notata von verschiedenen Aussenbedingungen. Z. vergl. Physiol., Abt. C, 22, 604-613.
ANIMAL BEHAVIOUR, 18, 3
484
Szlep, R. (1961). Developmental changes in the web spinning instinct of Uloboridae: Construction of the primary type web. Behaviour, 17, 60-70.
Tilquin, A. (1942). La Toile Géométrique des Araingnées. Paris: Presses Universitaires de France.
Witt, P. N. (1956). Die Wirkung von Substanzen auf den Netzbau der Spinne als biologischer Test. Berlin: Gottingen, Heidelberg: Springer.
Witt, P. N. (1963). Environment in relation to the
behavior of spiders. Arch, environm. Hlth> 7,4-12.
Witt, P. N. & Baum, R. (1960). Changes in orb webs of spiders during growth (Araneus diadematus Clerck and Neoscona vertebrata McCook). Behaviour, 16, 309-318.
Witt, P. N., Reed, C. F. & Peakall, D. B. (1968). A Spider’s Web. Heidelberg: Springer-Verlag.
{Received 12 June 1969; revised 20 December 1969;
MS. number: a856)