Images Collection



View this article in Search Friendly Plain Text
NOTE: This plain text article interpretation has been digitally created by OCR software to estimate the article text, to help both users and search engines find relevant article content. To read the actual article text, view or download the PDF above.
Experience and the Orb Web
CHARLES F. REED
Department of Psychology Temple University Philadelphia, Pennsylvania
PETER N. WITT MABEL B. SCARBORO
Division of Research North Carolina Department of Mental Health Raleigh, North Carolina
DAVID B. PEAKALL
Division of Biological Sciences Cornell University Ithaca, New York
Reed, Charles F., Witt, Peter N., Scarboro, Mabel B., and Peakall, David B. (1970). Experience and the Orb Web. DEVELOPMENTAL PSYCHOBIOLOGY, 3(4): 251-265. Cross spiders (AYaneus diadematus Cl.) were confined in tubes for between 3 weeks and 194 days, from the period preceding building of a first web, or as adults. The first webs built after release were invariably found to be significantly smaller (shorter thread and smaller catching area) than those of littermates raised in large frames. The ampullate glands of nonbuilders were found to incorporate 14 C alanine at a lower rate than those of regular web-builders. Pulling thread from glands repeatedly before release, or allowing animals to build several webs, brought speed of silk synthesis and size of webs to control values: this indicates that scarce silk supply is the cause for small webs after confinement. No differences in web size and pattern were found between mouth-fed and web-fed spiders, if animals were otherwise raised the same way. But adult spiders who had never been allowed to catch prey on the web disregarded flies in their webs. Differences in plasticity of web-building and prey-catching behavior are discussed.
web-building early experience innate behavior confinement orb web cross spider
Received for publication 20 July 1970 Developmental Psychobiology, 3(4):251-265 (1970) © 1970 by John Wiley & Sons, Inc.
251
252 REED, WITT, SCARBORO, AND PEAKALL
It has not seemed likely that the form of the orb-web is modified by the spider’s experience in building and using it. Even the first webs of cross spiders (Araneus diadematus Clerck) are miniatures of the adult form [Fig. 1 (a) and (b)], provided that the spiders hatch from their cocoons at the normal time (Peters, 1969). If confined in small tubes for several weeks after hatching, spiders upon release will build webs which seem to be indistinguishable from those ordinarily constructed. Petrusewiczowa (1938) and Mayer (1953), who performed such experiments, concluded that the behavioral patterns of web-building are innate mechanisms, maturing independently of practice. Given this evidence, it seems improbable that use of the web might be a reinforcement for web-building behavior.
Without denying the reasonableness and even the validity of this hypothesis of the innate nature of web-building, we will present evidence that the web is in fact not in its normal state after confinement of the spider for as short a period as 3 weeks. Webs built after 35 to 194 days of confinement (from hatching) are geometrically adequate, but show other effects of the treatment. Refinement of measurement and statistical comparisons have revealed differences not apparent in Petrusewiczowa’s and Mayer’s observations.
The stimulus to these experiments was the finding that Argiope aurantia Lucas who had spent their first 3 or 4 months outside the laboratory built adult webs in the laboratory which were larger than the webs of spiders raised indoors (Reed, Witt, & Scarboro, 1969). Two explanations among others occurred to us: (1) that emptying the silk glands through regular web-building stimulated silk production and (2) that successful use of the web as a trap reinforced building in the outdoor-reared group. The following experiments examined these hypotheses.
General Method
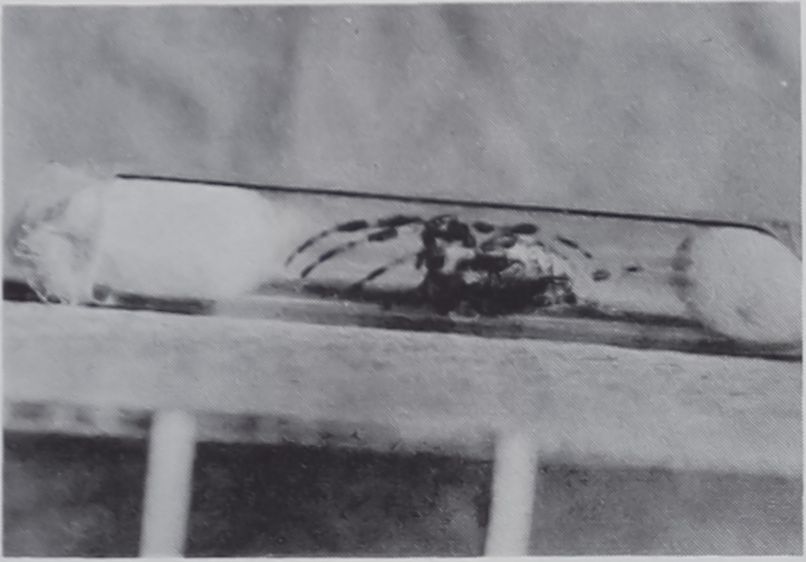
Spiderlings (Araneus diadematus Cl.) were confined, before they had begun web-building, in glass tubes of 3.5 (later 10.0) mm diameter and 65 mm length, with cotton stoppers at both ends (Fig. 2). The animals were unable to build webs in these tubes, although they were able to move about and to catch flies placed in the tube.
Littermates from the same cocoon were permitted to build in standard (50 X 50 X 9 cm) aluminum and glass laboratory frames, and were fed in the webs.
Each web was photographed the morning it was built. Calibration of these photographs and the deriving of web measures have been described in detail elsewhere (Witt, Reed, & Peakall, 1968). A representative set of web features was subjected to analysis of variance (3-factor, 2X2X2), with a repeated
EXPERIENCE AND THE ORB WEB 253

Fig. 1. (a) Sample web of a female Araneus diadematus Cl., 2 months old, as it was built ini the laboratory frame. The body weight of the individual Which had built this web was 7.8 mg, its leg length 6 mm. The 3 vertical bars in the upper right corner are spaced 10 mm apart, (b) Web built by the same spider at age 7 months; its body weight was now 139.9 mg, its leg length IS mm. Scale as in (a). Note upper end of small wooden frame in lower part of picture; such frames were placed into the 50 X 50 cm aluminum frames to provide the spider with a choice for web-building (compare with Fig. 4).
measure for each animal immediately after the confinement period and 1 week later (Winer, 1962, p. 337). Since some animals failed to build on that schedule, it was necessary to record more webs than were eventually analyzed.
Factors will be described in the details of method for each experiment. The set of web features was the same throughout, containing measures for’web size, density, and regularity: (1) thread length: total linear extent of all thread inside the frame of web; (2) spiral area: annular region covered by sticky spiral thread (the “catching zone”); (3) number of radii: all radial threads originating at hub; (4) number of spiral turns: a count of spiral threads in bottom portion of the web; (5) mesh width: spiral area divided by product of number of radii and number of spiral turns (a density measure); (6) standard error of median mesh size: the variation of a set of meshes (the area formed between adjacent radii and adjacent spirals) in the lower portion of the web; (7) angle regularity: variation in central angle size for adjacent pairs of radii.
Experiment I: Tube-rearing vs Unrestricted Web-Building Method –
Two groups of. spiders were kept in tubes for 7 weeks and for 28 weeks, respectively. At the time of release,- the animals confined for seven weeks did not differ significantly in weight from their littermate controls who had built freely

254
REED, WITT, SCARBORO, AND PEAKALL

Fig. 2. Adult female Araneus diadematus Cl. in glass tube with cotton stoppers at each end. View of animal is oblique from above: dorsal aspect of abdomen is visible as ovoid mass to the right, but cephalothorax is largely concealed by the 1st and 2nd legs on each side, which are arched upward and forward, to the left of the picture. The 3rd and 4th pairs of legs on each side are extended backwards. As may be apparent, the position of the animal was cramped. It could not build a web. It could, however, move about, catch flies and emit thread. Size is indicated by the 2 parallel rods of the scale shown below the tube; the rods are 20 mm apart.
in the same period. The mean weights were 8.9 and 10.8 mg, respectively. All 28-week-old animals were of course heavier than the 7-week animals, but the tube reared spiders at 28 weeks were much lighter than their free littermates (85 as opposed to 133 mg mean weights).
There were 5 experimental animals and 5 controls in each of the treatments, i.e., 20 spiders in all. Measurements were made on the first webs built after release date, and 1 week later. An average of 4 webs per animal had been built in the intervening period.
Results
In the following description, factor A refers to the condition of rearing—tube vs cage (or frame) controls, B refers to age of spider at release time, and C to the week interval between measures.
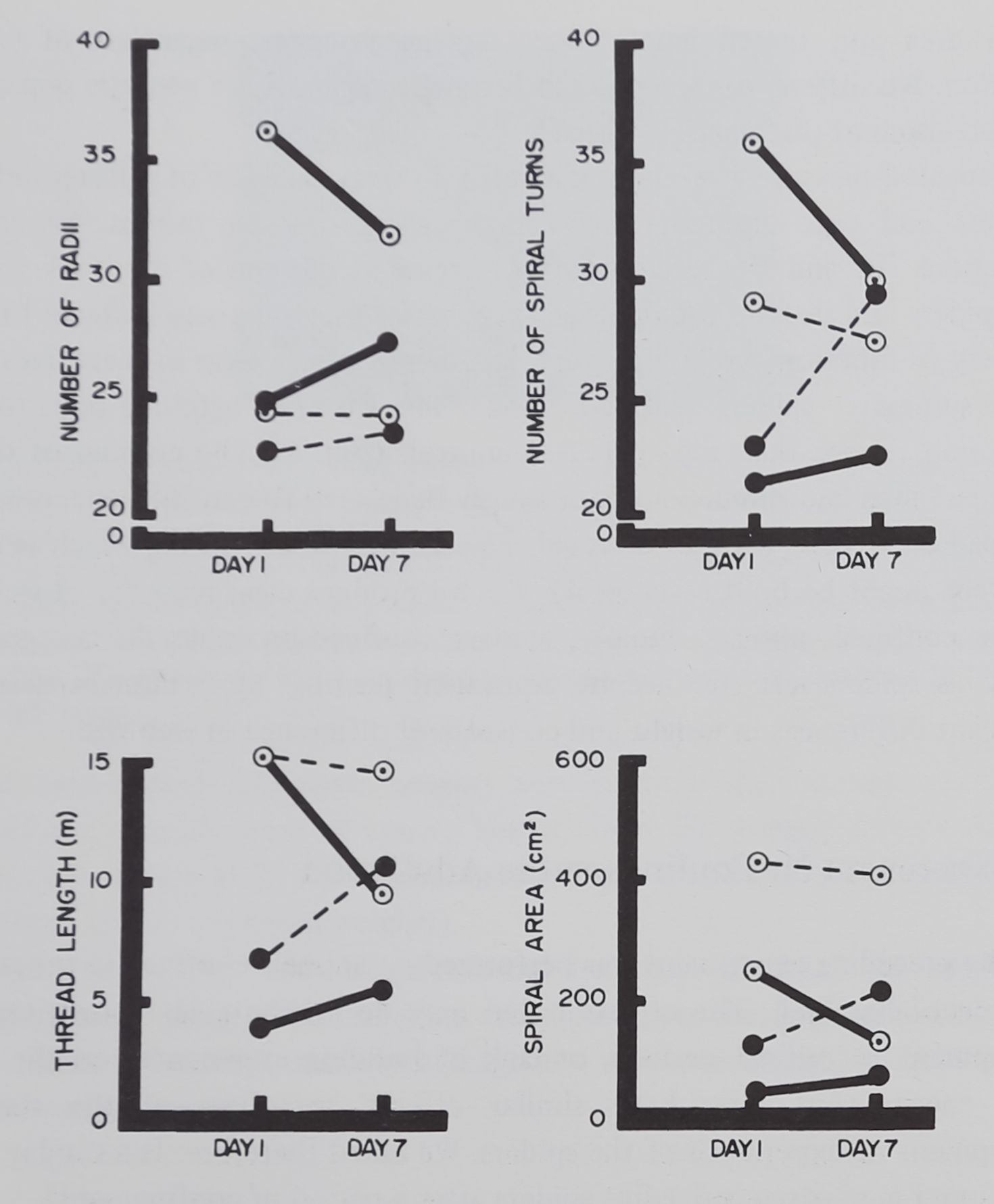
Rearing conditions (A) produce significant main effects in web size. For thread length (F = 94.75; df I 1, 16; p < .01) the means for first webs for tube and cage animals are 5.22 m and 15.20 m, respectively; for spiral area (F = 74.89; p < .01) the means are 81 cm2 and 345 cm2, respectively (Fig. 3). The related measures for number of radii and spiral turns differ for rearing groups, but there are significant interactions with age (B) that intervene: the young free spiders built many more radii and spirals than did any other groups. The density of threads (mesh measures) is accordingly also greater. This finding is consistent with earlier developmental data (Witt & Baum, 1960; Reed et al., 1969). Also, as would be expected, the older (and therefore heavier) spiders build webs of
EXPERIENCE AND THE ORB WEB 255
greater area and thread length than do the younger, regardless of rearing condition. No differences were found for angle regularity, a measure depending upon precision of placement of threads.
Repeated measure after a week yielded no clear evidence of different effects for tube and cage animals, or for that matter, for age groups. Significant interactions AC and BC resulted from increase at the end of the week for the tube spiders and decline for the caged; decrease for young and increase for old. Recovery of function, while suggested, was therefore not clear in these data.
In summary: spiders confined 2 and 7 months after hatching built smaller webs upon release than did their free controls (Fig. 4). The number of thread elements, radial and spiral, could not simply be related to confinement; precision of thread placement was not affected. Passage of a week, during which as many as 7 webs might be built (average 4), did not produce clear recovery of web size for the confined animals. Finally, spiders confined in tubes do not grow as rapidly as spiders left free despite equivalent feeding; by 7 months there are significant differences in weight and consequent difference in web size.
Experiment II: Confinement in Adulthood
The preceding experiment was performed on animals confined in tubes from the time of hatching. The effects noted may be attributable to restriction of development at critical periods, or lack of building experience; on the other hand, confinement may have similar effects regardless of the stage of development or experience of the spiders. We asked therefore: Is a similar effect to be found in experienced, adult spiders after a period of confinement?
Method
In this experiment, the webs of adult (28 week-old) spiders were measured before and after some of the group were confined in tubes for a period of 3 weeks. In contrast to the preceding experiment, the confined group had had normal and ample web-building experience before confinement.
Measurements were made of webs made immediately preceding and immediately following the confinement period.
Results
Because many of the confined spiders did not build webs upon being released, the analysis of variance presented here is based upon only 4 confined
256 REED, WITT, SCARBORO, AND PEAKALL
Fig. 3. Web features for short and long-term tube-reared and control spiders 1 and 7 days after release. Each point represents the mean for 6 animals; each line connects repeated measures on webs built by the the same animals. Solid circles: tube-reared; open circles: frame-reared controls. Solid line: 7-week group; dashed line: 28-week group. Note the significant difference between measures from webs built by spiders directly after confinement from those of all 3 other groups.
and 4 control spiders. Figure 5 contains summary data for the 2-factor analysis performed on the web measures.
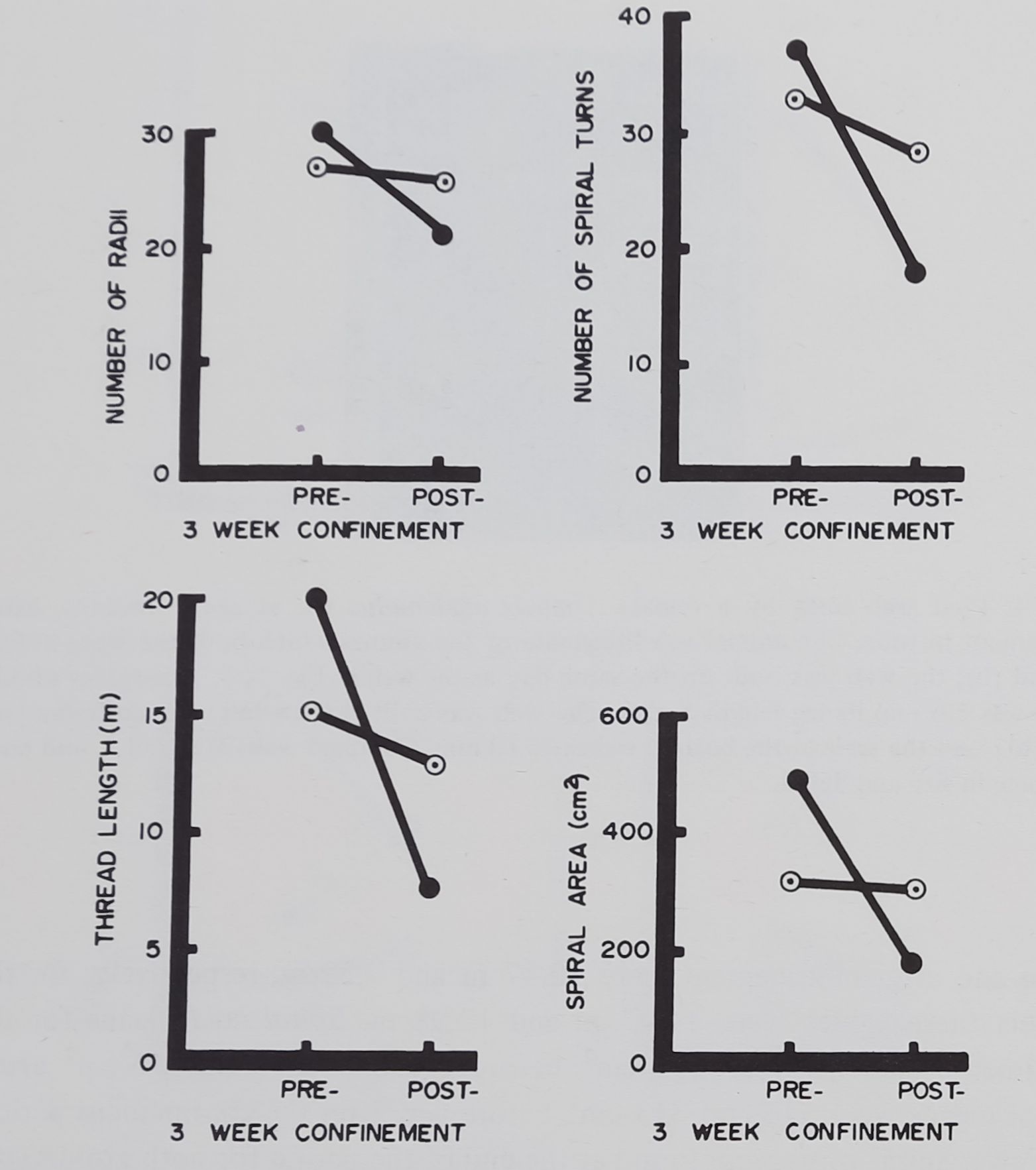
A significant interaction occurs between the factor of confinement and the factor of time for measures of thread length (F = 27.78) and spiral area {F = 29.92). (The critical value for .01 level for df= 1 and 16 is 13.75.) That is, there was little change in size of the web for the control animals during the 3-week period, while the post-confinement webs for the experimental animals were significantly smaller. The means for thread length for the experimental group

EXPERIENCE AND THE ORB WEB 257

Fig. 4. First web built by a female Araneus diadematus Cl. at age 2 months, after confinement in tube. The animal is a littermate of the animal which built the webs in Fig. 1(a) and (b); the web was built on the same day as the web in Fig. 1(a). This spider’s body weight was 6.6 mg, its leg length 5 mm. The web was built in the small wooden frame [see Fig. 1(b)] and the scale at the bottom indicates 10 mm. Compare webs 1(a) and 4, and note difference in size and detail.
before and after confinement were 20.47 m and 7.52 m, respectively; for the controls these values were 14.97 m and 12.98 m. Spiral area means for the experimental sample were 495 cm2 before confinement and 167 cm2 after; control means were 316 and 313 cm2 before and after the confinement period.
Fewer spiral turns were found at the end of the period for both groups (p < .01) and possibly fewer radii (p < .05); we interpret this shared effect as a developmental trend.
Regularity of thread placement was unaffected.
It may be concluded that confinement in adulthood produces web effects similar to those produced by confinement from hatching. The effect has to do with restriction of movement or web-building at any time of life, and not with a specific era.
The after-effect of confinement appears to be employment of less than a normal amount—at least a less than normal length—of thread in web construction. It is conceivable that the spider employs less than the full amount of material available in the glands, but the result also suggests an end-product effect; the presence of a certain supply of silk material in the gland may act to inhibit production of new silk. If this were the case, non-use of the material would eventually result in a lesser supply than that for glands regularly depleted in web-construction.
258 REED, WITT, SCARBORO, AND PEAKALL
Fig. 5. Web features for adult female spiders before and after confinement for 3 weeks. Each point represents the mean for 4 animals; each line connects repeated measures on webs built by the same animals. Solid circles: before and after confinement; open circles: unconfined controls. Note the relatively large difference in webs before and after confinement, and compare to insignificant changes in controls.
Experiment III: The Effect of Gland Depletion: Web Features Method
The role of gland-emptying and refilling was investigated by artificially depleting the glands of a sample group of tube-reared spiders. If an orb-weaver is pushed from a surface to which it has made a thread attachment, it lowers itself rapidly on the guy line which it emits from the spinnerets. By attaching the thread to an axle of a small motor, the thread may be wound as the spider

EXPERIENCE AND THE ORB WEB 259
releases it. This thread material comes from the ampullate glands; it is the silk used in the construction of the scaffolding of the web and possibly as a component of the thread for the sticky spiral.
The ampullate glands were emptied of silk for 2 successive days just prior to release for 6 spiders reared in tubes until 12 weeks old. Six frame-reared littermates served as controls. They built webs freely during the first 12 weeks of their lives. Building a web essentially empties the ampullate glands (Witt et al., 1968).
The animals were of equivalent weights. All procedures, apart from pulling, were as in Experiment I.
Results
The web measures of the animals reared in tubes do not differ significantly from the measures of their frame-reared littermates. The mean values for the first webs after the confinement period for tube and frame reared spiders are, respectively: thread length: 8.3 and 7.3 m, spiral area: 144 and 132 cm2; number of radii: 28.6 and 28.3 \ number of spiral turns: 26.5 and 21.0. As in the previous experiment, angle regularity did not differ between the groups.
Webs made a week later showed no differences between the “pulled” tube animals and their controls.
Artificially emptying the glands by pulling thread to depletion for 2 days brings the webs of the tube animals to the size of the webs of their free littermates. The typical result of confinement does not occur for the “pulled” spiders, consistent with the hypothesis of an end-product inhibition effect.
Experiment IV: The Effect of Gland Depletion: Production and Supply of Silk
The amount of silk material in the glands and its rate of production can be measured rather directly. If the end-product hypothesis is correct, animals who have been building regularly and have therefore been depleting the thread glands, should show both a higher rate of production and higher gland content of thread material than should nonbuilders. Further, pulling thread from the latter, or allowing them to build, should bring content and rate of production to the level of the builders, providing the animals are of equivalent body size.
EXPERIENCE AND THE ORB WEB
261
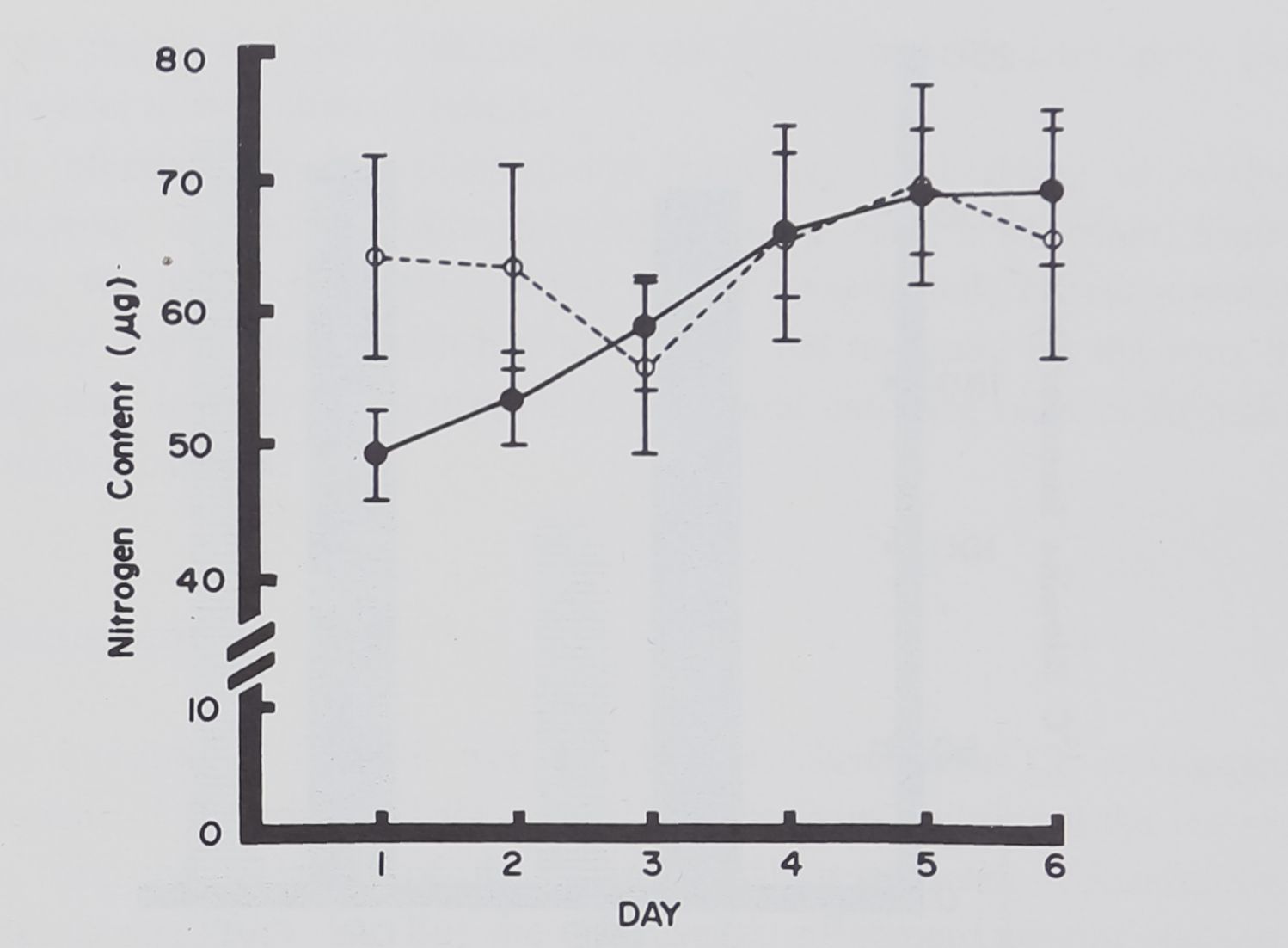
Fig. 6. Nitrogen content of daily pulled thread from spiders which had built webs for 2 weeks prior to Day 1, as compared with spiders which had not built during the same period. Open circles represent means of 6 builders, vertical lines standard errors; solid circles means of 10 nonbuilders. Note the differences between the 2 groups on Day 1 and 2 (significant at .05 and .01 level respectively) as compared to Days 3 to 6.
whether the utilization of the web, quite apart from its building, might have some facilatatory effect on subsequent building. The findings on the role of thread-exhaustion and replenishing do not bear necessarily upon this possibility. It is necessary to examine the form of webs built by animals who also utilize the webs for catching prey. Their subsequent web-building is compared with the web-building of spiders who built but did not use the web as a snare, and spiders who did not build but were fed in webs.
Method
Two groups of littermates (n = 18 in each group) were raised for 19 weeks in tubes and 22 weeks in frames, respectively. Half of the spiders in each group were fed on webs daily by placing a fly in the catching zone. The tube-reared spiders were placed in the vacated webs of other spiders for feeding. The other half in each group was fed directly, making the usual catching behaviors unnecessary: that is, flies were received in the tubes or brought directly into the grasp of the spider at the hub.
Thus the making of a web had various degrees of association with feeding: tube spiders fed in the tube and never on the web; tube spiders catching flies in

260 REED, WITT, SCARBORO, AND PEAKALL
Method
Ten spiders (Araneus diadematus) who had not built a web in the previous 2 weeks and 6 spiders who had built regularly in that period were confined in vials for a period of 6 days. Thread was pulled to exhaustion of the glands for all animals on each of the 6 days. Since silk is a well defined protein, the nitrogen proportion must be constant from sample to sample. Silk was digested in selenium sulfuric acid, and nitrogen content in the digested material determined with Nessler’s reagent in a spectrophotometer (Breed, Levine, Peakall, & Witt, 1964).
Results
Figure 6 graphs the daily mean values of nitrogen in the silk pulled from the animals. It indicates a steady progression from low values for the nonbuilders to the higher values of the spiders who had been building for 2 weeks. An analysis of variance treating the daily determination as repeated measures yields a significant interaction between group members and time. That is, the effects of pulling are different depending upon whether the animal had been building 2 weeks previous.
Apparently the effect of pulling has returned the nonbuilders to normal production level by the 3rd or 4th day.
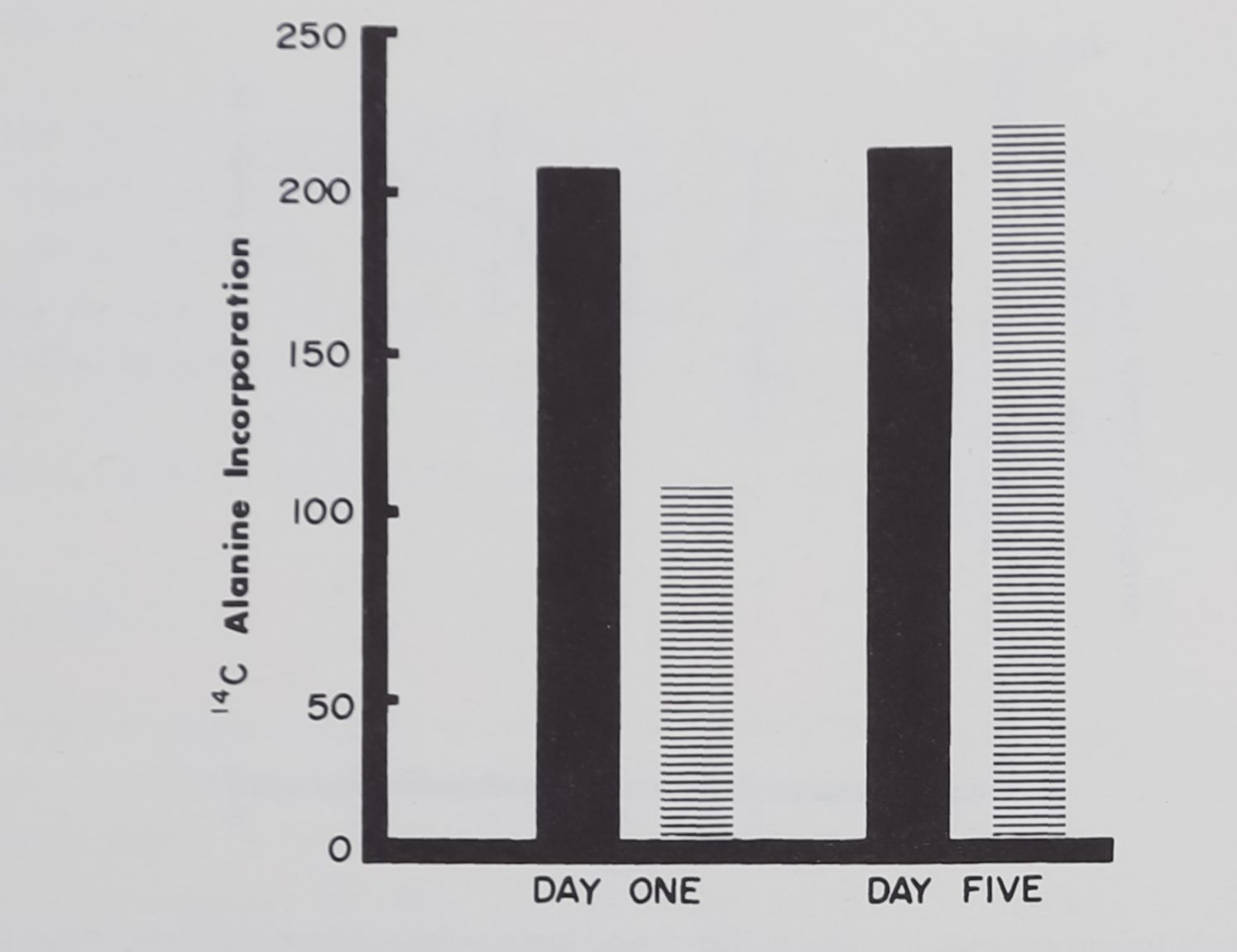
A corollary result is contained in Figure 7: it shows amount of incorporation of 14 C -alanine in a 4-hr period following emptying of the glands. The glands were dissected out of the spiders. Again nonbuilding and building spiders were the subjects of the investigation (n =?T2 for each group). Glands from 6 of each group were tested on the 1st day of removal of the glands. Glands for the remaining 6 spiders of each group were removed and tested after 5 days in which thread had been pulled from each spider daily.
The interaction between group membership and time of measurement is significant (.01 level). There are significant differences in incorporation between the builders and the nonbuilders on the 1st day tested; 5 days of pulling from non-web builders raised the synthesis rate to the level of the builders.
It may be concluded that pulling mimics the effects of depletion of thread through web-building: depletion leads to a progressively greater accumulation of thread material through increased rate of synthesis of silk. Clearly by Day 5 the gland is operating at normal levels.
Experiment V: Feeding and Web Size
To this point, the problem has been treated as if only a negative effect of confinement were involved. There remains the second hypothesis question
262
REED, WITT, SCARBORO, AND PEAKALL
Fig. 7. Four hours incorporation of 14C alanine into ampullate glands in vitro. On Day 1, 6 animals (black bar) had built webs for 2 weeks previously; 6 animals (hatched bar) had not built. On Day 5,12 animals, 6 of each group, had been pulled for the preceding 5 days, and both groups of 6 glands now incorporated alanine at about the same rate.
normal fashion on webs they had not made; spiders fed on webs they had made but in a manner which did not use the web as a snare; and spiders catching flies on their own webs in the usual manner. All spiders received the same number of flies.
The glands of the tube animals were emptied of thread material for 2 days before release, in keeping with the findings of the previous experiments.
Results
After 19 weeks the mean weight of the tube animals was half that of the free animals despite identical feeding schedules. Consequently, there are the usual differences in web size between the large unconfined animals and the small tube animals. Stimulating thread production by pulling increased glandular content of the small spider but did not bring it to the level of the large spider.
There were no differences in the webs of the spiders who employ the web in the normal fashion and those of spiders who had flies delivered directly to their

EXPERIENCE AND THE ORB WEB 263
grasp. As far as the data indicate, the use of the web to catch prey has no consequences in web characteristics.
No inference can be made about frequency of building as related to prey-catching, nor can sensitization or learning effects of other kinds be excluded. We noted, but have not yet further investigated, a most uncommon behavior of the animals which had never been fed normally on the web. They failed to turn toward prey struggling in the spiral catching zone or to make an effort toward capture.
Discussion
The building of the orb web by Araneus diadematm Cl. is affected by confinement of the spider. If the animal is kept from building webs for several months from the beginning of life, it fails to attain the mass of littermates and its webs are smaller in size. But the effects of confinement are not restricted to this condition. Spiders of equivalent weight at 7 weeks of age show the effects of being kept in tubes, as do spiders put in tubes for 3 weeks during adult life.
The effect is not an obvious derangement of the web, but a reduction in the area which it covers, and in the linear extent of the thread composing the web. Sometimes the number of radii and spiral turns is less than in controls, but this finding is not invariable. Statistical comparison of the webs of confined and control animals is necessary to demonstrate the effects Petrusewiczowa and Mayer, who did not employ these techniques, only observed—correctly—that the geometry of the web was not altered by confinement.
Although there appeared to be a trend for recovery of normal size of web during the course of a week of web-building after release, it is not clear that the effect of confinement has gone in a week.
Since the phenomenon is one of web size, it would seem to depend upon the supply of material in the silk glands, rather than upon any experience such as practice in building the web. Supply depends upon regular depletion, which in turn acts as a stimulus to synthesis of the proteins for silk material. Failure to deplete inhibits this synthesis: a form of end-product inhibition. In the case of the confined spiders, it can be supposed that insufficient emptying of silk glands had slowed synthesis to a level where only small amounts of silk would be available for the first webs built after release.
This hypothesis is supported by results of experiments in which the ampullate silk glands of confined animals were simply emptied for 2 days prior to release. Their webs were then of the size of control spiders who had been building freely-on an approximately daily basis. Further, by determination of the nitrogen content of the webs, it was shown that the amount of material in
264 REED, WITT, SCARBORO, AND PEAKALL
the glands of inactive spiders can be raised to the level of active spiders in a matter of 3 or 4 days. Similarly, the incorporation of material for synthesis could be accelerated in previously inactive spiders; this change was accomplished by depleting silk glands over a 5-day period, as shown by the data on incorporation of14 C alanine in Experiment IV.
Ordinarily, building a web empties the ampullate glands of silk (Witt et al., 1968), as does the thread-pulling procedure. The biochemical data seemed to indicate that the supply of silk in inactive spiders could be brought to control levels in less than a week. The data on web size, however, did not clearly indicate that control values had been reached a week after release from the tubes. To the extent that web size is dependent upon silk supply, these results are not quite in accord. Two methodological points should be noted: pulling occurred on successive days while web-building did not, and confinement times were shorter in the biochemical experiments. After 5 days, for instance, glands had not necessarily been emptied by building 5 times.
It is concluded that the achievement of an age and size-appropriate web, in size and in pattern, does not depend upon experience in building. Size is affected by confinement, but the significant variable is supply of thread. Petrusewiczowa and Mayer failed to observe this phenomenon, but their conclusion that web-building is unaffected by experience must be allowed to stand.
It is important to note that web regularity and shape were not affected in the smaller webs. The spider with reduced supply of thread may be said to adapt to this condition. It does not run out of material somewhere in the middle of construction. Nevertheless, it need not be supposed that this adaptation is acquired by experience or that building techniques have to be perfected.
This experiment does not lend support to the hypothesis (Reed et al., 1969) that use of the web in catching prey has an effect upon the size of the web or—in any case less likely—upon its pattern. Nor is;the successful employment of the web a positive reinforcement to web-building, as far as our data indicate. A spider who always had prey delivered directly into its grasp (rather than having to respond to the stimuli produced by prey striking the catching zone of the web) seemed unresponsive to those stimuli when first encountered.
Building behavior itself does not seem a fruitful ground for investigating plasticity in the spider nervous system. Prey-catching may require some adaptation of the animal, although even here the web may be viewed as an achievement which has reduced the contingencies with which the spider must be prepared to deal. The web filters the prey population to appropriately-sized insects: those too large and too small escape.
These conclusions do not rule out the possibility that other forms of experience may serve to modify the behavior of the spider through habituation or through learning. Szlep (1964) has produced habituation to repeated
EXPERIENCE AND THE ORB WEB 265
vibratory stimuli; Bays (1962) has observed discrimination of tones related to palatable and unpalatable booty; and LeGuelte (1969) has demonstrated what appears to be learning of re-orientation in the inverted web. These are behaviors associated, not with web-building but with approach, avoidance and successful retreat; they may be more responsive to contingencies than is behavior involved in building the web. Looking at the problem from the standpoint of natural selection, web-patterns may be optimal and not to be improved through experience; prey catching in the web, on the other hand, has in some way to be adapted to the prey-population which a specific area provides at a certain time.
Notes
Supported by National Science Foundation grant number GB-6246X1 to Peter N. Witt and National Science Foundation grant number GB-5736 to David B. Peakall.
References
Bays, S. M. (1962). A study of the training possibilities of Araneus diadematus Cl. Experientia, 18: 423.
Breed, A. L., Levine, V. D., Peakall, D. B., and Witt, P. N. (1964). The fate of the intact orb web of the spider Araneus diadematus Cl. Behaviour, 23: 43-60.
LeGuelte, L. (1969). Learning in spiders. Amer. Zool., 9: 145-152.
Mayer, G. (1953). Untersuchungen über Herstellung and Struktur des Radnetzes von Aranea diadema und Zilla X-notata mit besonderer Berücksichtingung des \Unterschiedes von Jugend und Altersnetzen. Z. Tierpsychol., 9: 337-362.
Peters, H. M. (1969). Maturing and coordination of web-building activity. A mer. Zoologist, 9: 223-228.
Petrusewiczowa, R. (1938). Beobachtungen über den Bau des Netzes der Kreuzspinne {Aranea diadema L.). Trav. Soc. Sei. Lettr., 13: 1-24.
Reed, C. F., Witt, P. N., and Scarboro, M. B. (1969). The orb web during the life of Argiope aurantia (Lucas). Develop. Psychobiol., 2: 120-129.
Szlep, R. (1964). Change in the response of spiders to repeated web vibrations. Behaviour, 23: 203-230.
Winer, B. S. (1962). Statistical Principles in Experimental Design. New York: McGraw-Hill.
Witt, P. N., and Baum, R. (1960). Changes in orb webs of spiders during growth (.Araneus diadematus Clerck and Neoscona vertebrata McCook). Behaviour, 16: 309-318.
Witt, P. N., Reed, C. F., and Peakall, D. B. (1968). A Spider’s Web. Berlin: Springer Verlag.