Images Collection







View this article in Search Friendly Plain Text
NOTE: This plain text article interpretation has been digitally created by OCR software to estimate the article text, to help both users and search engines find relevant article content. To read the actual article text, view or download the PDF above.
Spider Webs:
Design and Engineering
INTERDISCIPLINARY SCIENCE REVIEWS
J. WESLEY BURGESS and DR PETER N. WITT
Research Section, Mental Health Division, Department of Human Resources, State of North Carolina, U.S.A.
‘All things in nature,’ said Sullivan, ‘have a shape,
that is to say, a form, an outward semblance, that tells
us what they are, that distinguishes them from our-
selves and from each other‘Whether it be the
sweeping eagle in his flight or the open apple-
blossom, the toiling work-horse, the blithe swan, the
branching oaks, the drifting clouds, over all the
coursing sun, form ever follows function, and this is
the law.’
Joedicke, 1959
INTRODUCTION
The interdisciplinary approach of combining
architectural and engineering knowledge with biolog-
ical and ethological thinking can elucidate the reasons
for the enormous variety in spider webs; and web
analysis can teach us something about optimal forms
for the construction of a functional building.
When a house is designed by an architect, various
needs are taken into consideration. First, the site will
always interact with the structure; for example,
environmental conditions may demand a strong, or
permit a more open design. At the same time, the
building becomes an integral part of the environment,
changing it in a way which may influence future
building nearby. It makes a great difference whether
structures are meant to stand over long periods of
time, or whether they serve temporary needs, to be
taken down after a few days.
Ecofiomy of material and labor influence the way in
which a building is planned, and available material
dictates the width, height and shape of rooms. Human
buildings show wide variety in size, depending on
whether they are meant to be inhabited by one indi-
vidual, a single family, or a great number of people. If
they have mainly a sheltering function, their form
must be different from office and industrial structures.
In addition, separate units are connected with each
other by transportation networks of highways and
railroads.
Spiders live in self-made structures too. There is an
enormous variety in spider webs which is noticeable
even to the casual observer. Near the ground there
may be webs which contain dense tangles of threads
or silken sheets; in high grass and in shrubs there are
more elaborate three-dimensional structures, while
up in the trees beautiful geometric orbs can be recog-
nized. Some house only a single animal, others shelter
a mother and her offspring, and still others are large
communal complexes inhabited by hundreds of indi-
viduals.
A great variety of roles has been suggested for
spider webs: some of the better known are network
for movement and communication, silken trap, pro-
tective device, and platform for mating. In a number
of instances we have experimental evidence for the
part played by a structural detail, in others the func-
tion can only be surmised. In specific webs, the sticky
catching thread is neatly separated from the dry silk
of the framework, setting the catching function apart
from that of support. Signal threads run unimpeded
through open spaces to allow undampened transmis-
sion of information.
The resonant properties of webs filter out inap-
propriate signals, and specific thread patterns send
information in many directions, so that a multitude of
animals can be recruited for an attack on oversized
prey. Orbs and tangles can be intermingled to form a
composite functional habitat for a small group of
spiders. Broad opaque silken bands protect builders
from the eyes of predators. And the communal tangle
built by some baby spiders creates a favorable envi-
ronment for growing up.
We postulate that each structural feature in a web
can be understood as part of a system of strategy for
survival in the builder’s own unique environment. We
believe that the comparison of spider structures with
human buildings (Fig. 1) will identify basic principles
in design which serve similar purposes in two groups
of otherwise dissimilar living beings, revealing the
systems by which both modify their environment.
In the drawing opposite, Edward H. Williams, a Raleigh, North Carolina architect, superimposed various building
designs discussed in the review: the floor plan of a family residence with side terraces, at bottom a suspension
bridge, a clover leaf highway interchange at right, and the symmetric orb web of a spider, left top; all suspended in
an irregular space structure. Each of these designs shows a solution to a problem in the builders’ lives which is
examined in the text of the review.
Figure 1. In this 16th/17th century Dutch print, a group of people consider howto lay out a city
while they look at a spider web. The symmetric orb design of the web is reflected in many city
plans, like the Place de l’Etoile in Paris, orthe radial plan for Karlsruhe where the main streets begin
at the centrally located castle of the prince. Such city plans permit the ruler to keep the citizens
under control by positioning guns at the center, comparable to a spinder’s control over prey in the
web through the vibrating radii.
THE BUILDING SITE
In human affairs we are apt to consider man as a
self-sufficient unit. Actually, people are not
autonomous, but with their organizations and hous-
ing structures they form a total system from which no
part can be separated. How complete is a man alone
without his society or country? Similarly, a farm
in the absence of the farmer, his family or the live-
stock does not function. In the same way, it is hard to
talk about a spider without a web or the web without a
spider.
The blind spider without a web is nearly helpless,
unable to catch or even identify prey without vibra-
tion cues (Baltzer, 1923), and his hooked feet,
adapted for hanging on silken threads, slip on smooth
surfaces. Conversely a naked web will not catch or
hold prey and quickly falls into disrepair. An even
DR PETER 1NL WITT studied medicine in Berlin and Graz and obtained his medical degree
from the University of Tübingen. He taught pharmacology at the Universities of Bern,
Switzerland, Harvard Medical School and State University of New York at Syracuse. For the
last 10 years he has directed mental health research for the State of North Carolina and held
faculty appointments at N. C. State University and University of North Carolina at Chapel Hill,
N. C. His interest in spider webs began with the discovery that drugs alter web patterns in
1949. Since then he has published widely on web building under various normal and stress
conditions. For the last two years he has worked with J. Wesley Burgess on webs and spiders
under a grant from the National Science Foundation.
J. WESLEY BURGESS studied entomology and zoology at Purdue and North Carolina State
University. During field trips he collected social and parasocial spiders, and developed
methods for evaluating their behavior.
Address: Research Section, N.C. Mental Health Division, Box 7532, Raleigh, North Carolina
27611, U.S.A.
doser tie exists between spider and web: the silk is
secreted by the body and eaten by some species in
hard times to become part of the body again. In fact,
orb weavers eat their webs regularly and secrete 95%
of the same materials the next day (Peakall, 1971),
simply transferring amino acids in and out of the body
for utility’s sake.
For these reasons we will think of web and spider
(and architecture in general) as an integrated system.
As such a complex, the web/spider unit can survive; it
enters into the global exchange of energy and materi-
als, and this is what biologists study. Products can be
transferred within the complex, as when orb web
building materials are secreted and reingested.
At the interface of the web/spider organism there
are interactions with the environment, such as taking
in food and oxygen and releasing carbon dioxide and
faeces (Fig. 2). Again, some web/spider complexes
might be likened to a farm, where an integration of
sub-units, like cow, grass and farmer, function
together. Here also, there can be intraexchange wBn
the farmer milks the cow or spreads manure on the
meadow. Interexchange occurs with the outer envi-
ronment when the farmer buys hay for the cow or sells
the milk. In fact, we are familiar with many systems
which are like this.
A human city, for example, may provide Its own
services, such as garbage collection and fire protec-
tion, or generate its own electricity; interactions can
occur involving transfer of energy and materials
within the ‘city/citizens’ complex, pot food or tele-
phone service a city must rely on interexchange from
outside its boundaries. If we integrate the city fun|S
tions with a huge architectural structure, as proposed
by Paolo Solari, we have an attractively close parallel
to a spider colony. Still, in such a system no unit is self
sufficient. Fires, trucks, buildings, food and firemen
are interlocking parts in a complex which is successful
in its ability to sustain itself and to replace its compo-
nents over time.
With both the web/ spider and the farm and city,
however, it is not enough simply to bring the several
sub-units together: success of the structure depends
upon finding a good place to operate—a proper
potential site. When ecologists talk of ‘site selection’
or ‘site utilization’ in spiders and other animals they
are generally considering site to be an area of material
resources. However, we prefer to look at the broader
spectrum of interplay between an organism and its
environment (Fig. 2).
In this way we avoid difficulty in discussing the
web/spider complex, because the spider can set up
housekeeping literally in thin air. G. C. Argon (1957)
states this as a concept of design, that ‘By joining with
uninterrupted lines a number of points in space, the
object claims a “site”, and existence in undetermined
space’. From the design view, all that we investigate in
webs is a product of their creation in defining a site,
and this has no relation with the formerly uncolonized
area, except as a source in space in which to put a new
object: the web. This is because all the site require-
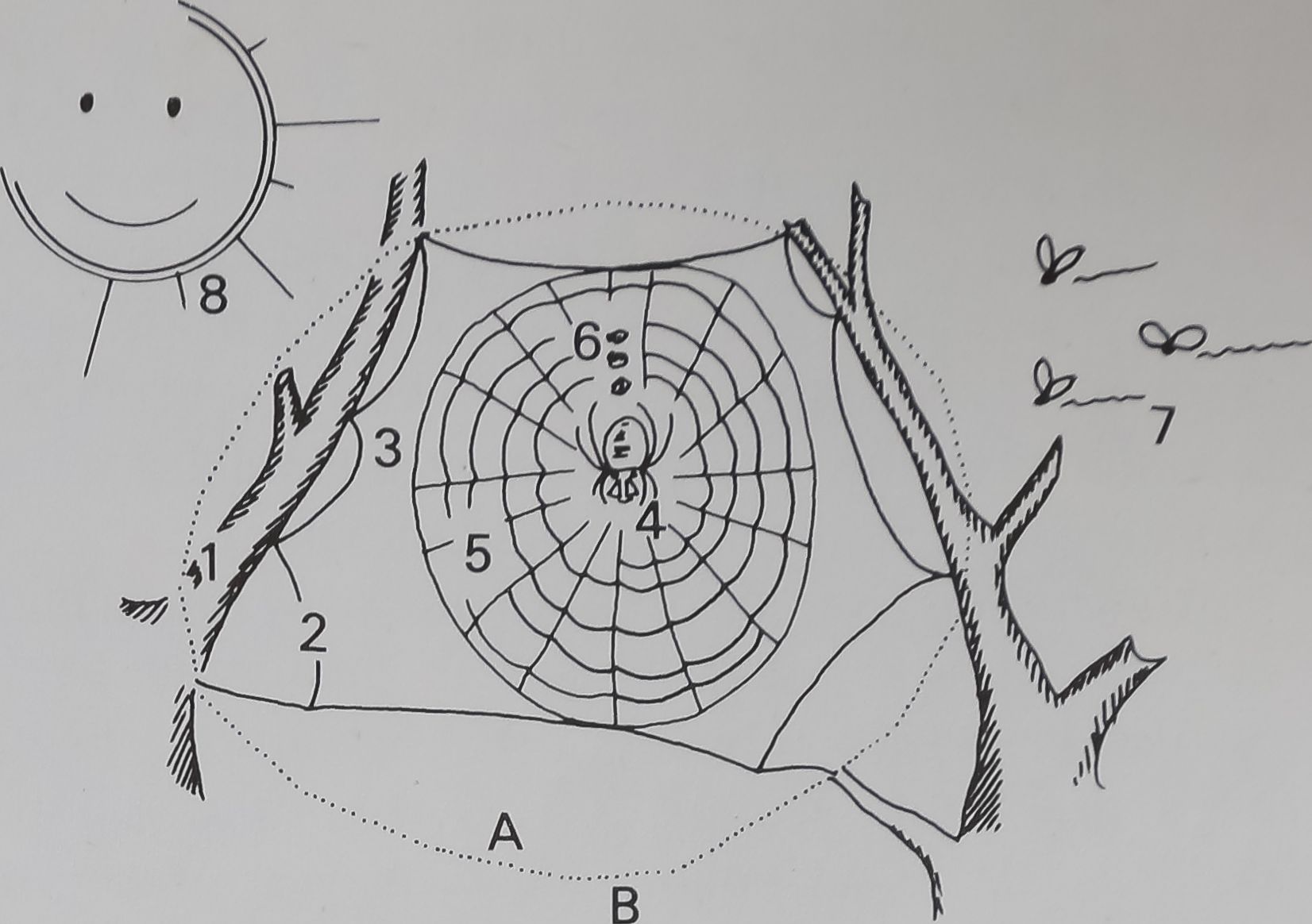
Figure 2. The site area defined by web/spider
complex of Araneus diadematus. A. Site area,
also web space; B. Non-site area or non-web
space. Web/spider complex components: 1.
Environmental structural components,
nature’s contribution to web site. Contribute
potential and kinetic energy to complex. 2.
Buffer zone, cushions and filters exchange
with non-web space. 3. Web sector, perma-
nent structural part, potential store of
metabolic energy. 4. Spider sector, living part,
potential store of metabolic energy. Constant
material loss from body processes. 5. Web
sector, temporary structural part, potential
store of metabolic energy. 6. Stored prey,
store of metabolic energy. Some loss through
degradation. 7. Prey source, metabolic and
kinetic energy input. Material input. 8.
Environmental interface, source of material
(water) input, heat energy, cooling forces,
kinetic energy (wind) and other diverse inputs
and losses.
The organization by the spider of previously
existing and internally produced components
into the site defines site (A) and non-site (B)
areas. We call the material part of this organi-
zation a structure, which is an important part
of the web/spider complex system. The
web/spider complex system filters and
buffers exchange with the non-site area,
allowing necessary supplies to come in and
wastes to pass out. Within the complex, the
temporary web is built within a permanent
structure, work is performed, energy is used
and stored. This general model may be
applied to other webs or other structures such
as a human office building or bank.
ments of any web/spider complex, whether structural
(strong supports, close twigs, open spaces in vegeta-
tion) or biological (rain water, sunshine, prey sources)
only become functionally available with the establish-
ment of the structure itself.
A corollary is that the first step in web system
strategy is to modify the immediate environment.
Other species may rely on morphological or
physiological adaptation for survival, but spiders and
man primarily adapt behaviorally by keeping their
physical form and changing their surroundings.
Environmental modification can come about through
the active choice of the spider (Turnbull, 1973;
Enders, 1974), or natural selection operating on large
INTERDISCIPLINARY SCIENCE REVIEWS, VOL. 1, NO. A, 1976 325
numbers of randomly placed offspring. One female
Araneus diadematus can produce up to 1000 spider-
lings, of these the few surviving to adulthood are
found in predictable foliage and positions. They may
have actively selected these spots, or have distributed
randomly when only the well-placed would survive.
In either case, the method is inconsequential for our
consideration, as the result is the same.
Araneus diadematus orbs are primarily found high
in trees and shrubs, and the same is true for the sheets
of Mallos gregalis (Diguet, 1915), while Cyrtophora
webs are found within 200 cm from the ground
(Wiehle, 1928) on opuntiae and agaves. Metepeira
webs are characteristically within a leafless open
space in a bush or tree (McCook, 1889). Whatever
the place or method of selection, the end result is to
delimit characteristic ‘web’ and ‘not-web’ areas for
each species. It is easy to imagine global creation of
diverse sites, so that all heights, hosts and geographies
are utilized with species living side by side. This is
exactly what we see when we look at nature: these
beautifully interlocking systems complement each
other, allowing all ‘niches’ to be filled and every
species to get its share of the biological pie. A com-
parison can be made with the view of the Earth
presented to the passenger in an aeroplane: the land-
scape appears neatly divided into farms, cities, for-
ests, and roads uniting various places. Such parallels
will grow increasingly obvious as we consider the
spiders’ complex in more detail.
THE SPIDER’S WEB
Components
For web components the following terms will be used
in this review. The geometric orb denotes a two-
dimensional cartwheel structure. It consists of radial
spokes around a hub; the radii are crossed by spiral
turns, and end peripherally in a complex frame. The
area covered by the spiral structure is frequently
elliptic rather than circular and the hub usually lies
off-center. All known orbs are short term structures,
and are repaired or renewed after a few days. Build-
ing time is short: one to two to a few hours. There are
two different procedures for constructing the geomet-
ric orb:
In the vertical Araneus type web, which we will
discuss in detail, all or nearly all radii are built first,
the majority running all the way from hub to frame;
later the spiral is laid across the completed radius
structure from the outside in (Fig. 3). Under the
microscope most thread crossovers look like inte-
grally fused junctions. The meshes of the orb are
rectangular. In Araneus webs, spiral threads are
covered with drops of glue, whereas radii are dry
(Witt, et aL, 1968).
Figure 3. This geometric orb web of an adult
female Araneus diadematus, or cross-spider,
was built in a laboratory frame in about 30
minutes. The builder sits on the hub and pulls
radii tight with its eight legs. A scale in the
upper left corner indicates 20 mm in the origi-
nal, and the direction of gravity, showing this
to be a vertical orb web. A small wooden
structure was deliberately introduced into the
lower right part of the larger frame: the spider,
which was prevented from leaving, has dis-
tinctly modified the spiral where it would have
overlapped the frame.
In the Cyrtophora web, which is a compound struc-
ture, a horizontal geometric orb is only one compo-
nent. When the orb is built, only a few radii (10-20)
are laid at first, then additional radii are constructed
as the spiral is laid from the hub outward, to a total
radius number of 300 to 500 at the frame. This
construction sequence is clearly explained by Kull-
mann (1958). Because radius and spiral threads run
together briefly, the meshes have a hexagonal shape
(Fig. 4E).
A sheet web is a two-dimensional structure, show-
ing no symmetry. It either lies flat or is bowl-shaped,
composed of long threads which frequently cross each
other unfused. There can be oval holes in the sheet,
which are lined with reinforcing thread, but otherwise
the structure is uniform without subdivisions. All
sheet webs we know forai long-lasting structures,
built intermittently over long periods of time.
The space web is three-dimensional, occurring
either by itself or together with an orb or sheet. There
are no clearly delineated substructures in a space
web; however, during construction great care is taken
that tension is evenly distributed in all directions
(Holzapfel, 1933). Space webs show some woven
parts, built with short strands which frequently are
fused into Y structures; other sections of the space
Figure 4. Web of Cyrtophora cit-
ricola, modified from Kullmann
(1958). A shows the upper space
web, which increases in density
toward the center; B the lower
space web, which is relatively
wide-spaced on top. In the middle
the bowl-shaped horizontal sym-
metric orb web C separates the
upper from the lower part, pro-
vides running space for the spider
at the underside, and acts as a
receptacle for falling prey. D pro-
vides a look onto the orb from
above, showing the increase in
radial number toward the
periphery, here at the bottom; E
shows an enlarged mesh of the
orb with its hexagonal shape.
web have wide open spaces, crossed by only a few
silken lines. Space webs are usually constructed in the
course of several days; the same web is used for weeks
and months, sometimes throughout the life of the
builder. Many space webs function as sites for com-
munal life and cooperative prey-catching.
One or more of the above components appear in
each of the four webs we selected, and in most other
known webs. We do not examine the widely distri-
buted single sheet webs of the Lyniphiid spiders,
which are obvious on a dew-covered meadow; nor do
we discuss the single-thread ‘web’ which the bola
spider keeps in perpetual motion.
Web Examples
We discuss four types of web. Firstly we have the
Araneus web (Fig. 3), typical of the many spider
species which construct single verticle orb webs. All
these webs are slight variations on the basic theme: a
radial support structure, overlaid by a so-called
catching spiral. The finished structure is particularly
beautiful. There is a logarithmic decrease of inter-
spiral distances from the periphery towards the
center, and usually the radial threads are spaced at
wider angles at the top than at the bottom. The spiral
area is oval, with the long axis vertical; and there are
circular as well as pendulum turns. In the middle are
the hub and open zone; outside is the irregular frame
area.
The frame is suspended on anchoring lines that
form a buffer zone between non-spider built struc-
tural elements, like branches and walls, and the spiral
area, which can have a number of individual varia-
tions. The single occupant sits either in the hub, or in a
hiding place outside the web, holding a signal
thread stretched to the hub. The web of Araneus
diadematus Cl. (commonly called the ‘cross spider’)
has been most thoroughly explored and recorded (for
a recent review of Araneus web literature see Witt, et
a/., 1968).
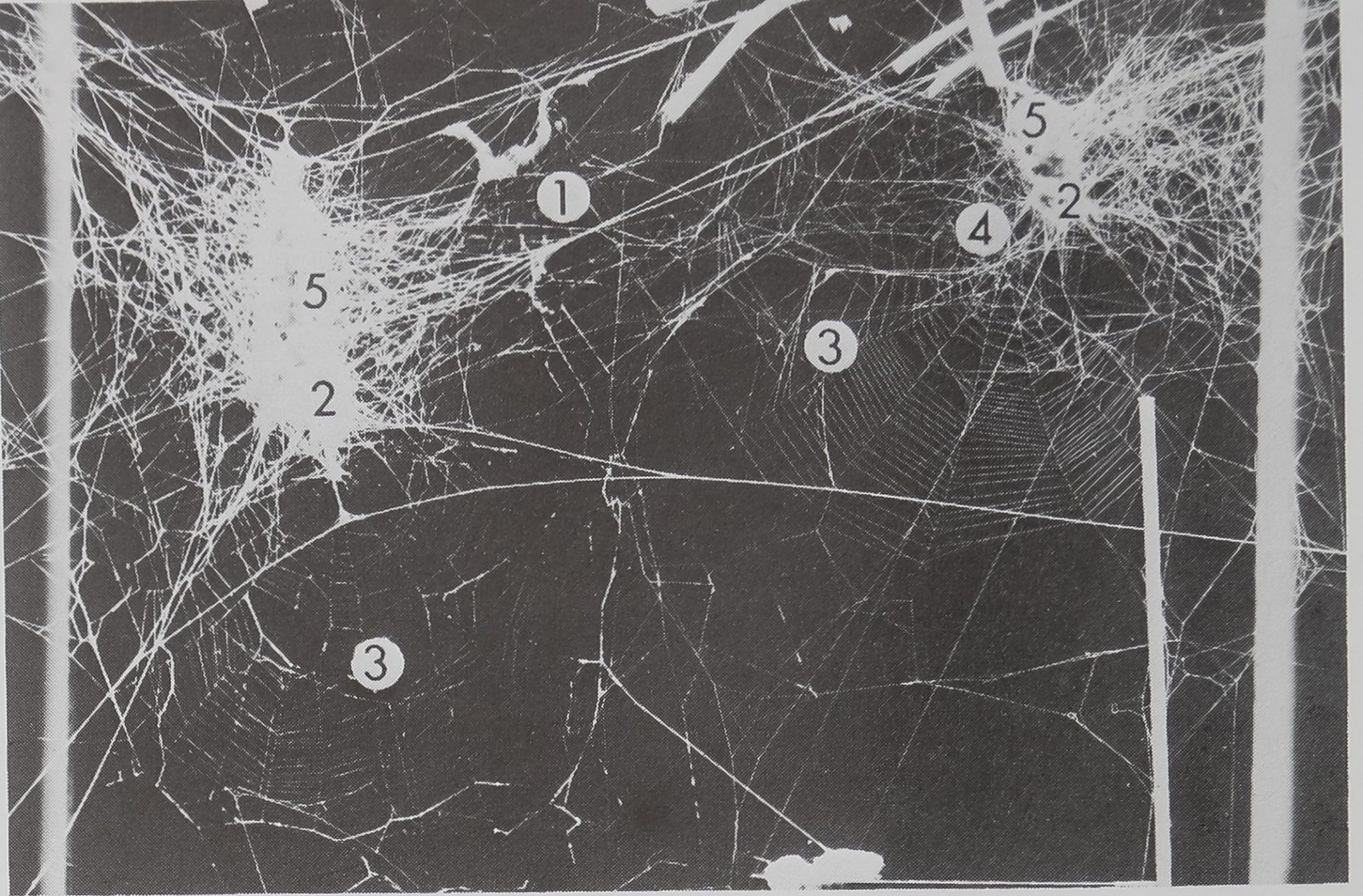
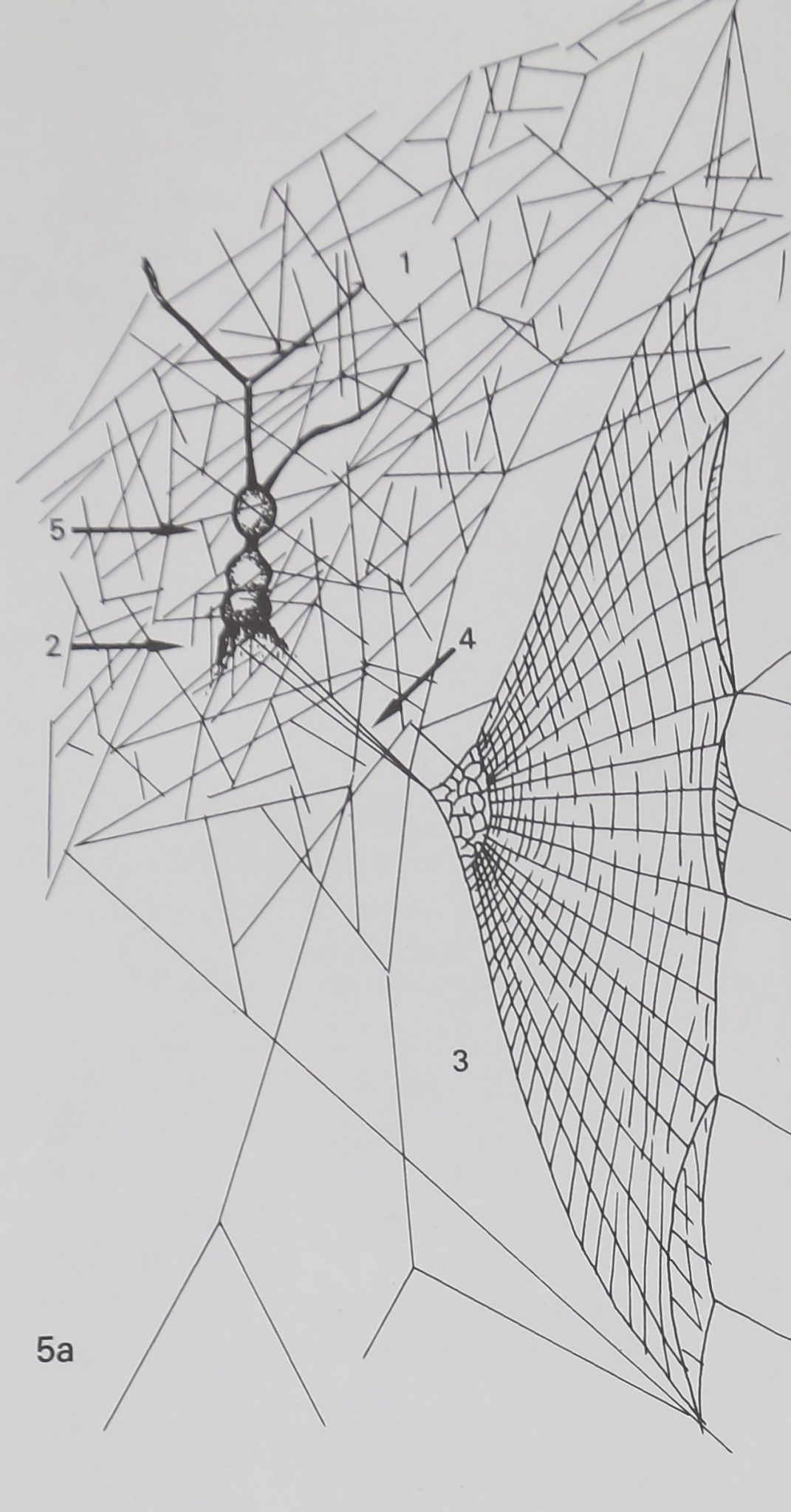
Then there is the Metepeira web (Fig. 5a and b), a
composite web built by members of the genus
Metepeira which is distributed world-wide. Two
species in particular, M. labyrinthea and M. spinipes,
show aggregations and a repertoire of interactions
(McCook, 1889; Pickard-Cambridge, 1903). The
web contains five distinct structural features, each of
which has different physical properties as a result of
different combinations of silk from the silk glands and
thread laying behavior, These features are space-
web, retreat, orb web, signal threads and egg sacs.
Each is distinct and all are built in a fixed sequence.
The orb component in Metepeira and Araneus has
the same structure. Numerous Metepeira may inhabit
a conglomerate of many webs, each animal building
its own retreat and orb.
Next we have the Cyrtophora web (Fig. 4); the
webs of Cyrtophora citricola and C. moluccensis
appear the same, and have been described and
analyzed by Wiehle (1928), Kullmann (1958), Blanke
(1972), and Lubin (1973). They are three-
dimensional structures, consisting of at least three
easily distinguishable features: a space web on top
and below, separated by a horizontal orb. The upper
part is a three-dimensional, irregular mesh-work.
INTERDISCIPLINARY SCIENCE REVIEWS, VOL 1, NO. 4, 1976 327
Figure 5a. One Metepeira compound web.Eiirst built is space web (1), where spider sits under
retreat (2), separated from orb web (3), and holds connecting threads (4), above retreat; mature
females may construct tiers of egg sacs (5). Woodcut by McCook 1889. These single web modules
may be joined together for colonial living.
Figure 5b. On the left and right are retreat/egg sac combinations containing Metepeira spinipes
individuals who have built webs sharing thread connections. Interactions occur in the permanent
connected space web, while prey is caught by individuals in periodically built orb webs. In nature,
many M. spinipes build webs together, sharing a single site. For explanation of numbers see Fig.
5a.
becoming increasingly dense toward the orb. A hori-
zontal orb forms the middle part of the web and, in
contrast to those of Araneus and Metepeira, has a
great number of incomplete radii. The flat orb is
distorted by strong vertical lines, so that it resembles a
bowl with a raised center. The area directly under the
orb is relatively open, permitting the single inhabitant
to move around rapidly. Often many webs are built
together (e.g., 200; Lubin, 1973) forming a colony in
which the individual structures are connected
through their space-webs and common mooring lines.
In these large communities hubs maintain a minimal
distance of 15 cm, and animals frequently exchange
webs.
Finally there is the Mallos gregalis web (Fig. 6).
These tiny spiders from central Mexico live together
in the thousands. The colonies consist of a single,
huge web on which all members coexist socially,
hunting prey and feeding in groups. The web as a
whole has the appearance of a grey mass, the newest
part being pure white. Again the three-dimensional
structure has three distinct features: the outermost
surface or prey-catching sheet, usually studded with
dead flies; the complex interior space web,
honeycombed with tunnels; the internal chambers
containing spiders and egg sacs. Like many other
spiders forming complex societies, Mallos gregalis
is prevalent in the tropics (Burgess, 1976), where
webs are constructed far above the ground in
trees or bushes. They sire easy spiders to keep in
the laboratory for they will accept a wide variety
of supports (Diguet, 1915; Gertsch, 1949; Burgess,
1976).
Web Boundaries
These sites need not be thought of as static or iso-
lated, any more than we consider a town as only an
insular unchanging structure. On the contrary, they
are transformed into a dynamic web/spider complex,
capable of exchanging energy internally as well as
328 INTERDISCIPLINARY SCIENCE REVIEWS, VOL, 1, NO. 4, 1976
Figure 6. Section of a tree colony of Mallos
gregalis observed near Guadalajara, Mexico.
Spiders communally build webbing around
branches and leaves; flying prey adheres to
the sticky exterior sheet where communal
feeding takes place. Spiders normally rest
within the interior of the web structure. Note
the openings in the covering sheet web, which
give animals access to the web surface.
externally with their environment. Araneus
diadematus builds a bridge thread and anchoring
lines between branches or twigs. This defines the
periphery of her solitary site, and also forms the
buffer zone between non-spider built supports.
Branches which were separate and formerly
undefined now, as anchor points, become a dynamic
part of the site system. The frame threads, compara-
ble to the foundation of a house, modify the shape of
these environmental supports to best suit the daily
construction of an orb web; they also absorb kinetic
stresses, forming a tension skeleton on which the orb
hangs.
This system is reminiscent of Klee’s sculpture: Tn
this structure, the dual factor of a system of suspen-
sion and a system of support is certainly apparent, but-
the oblique planes have full freedom within the sup-
porting framework to which are attached joints so
delicate as to be mere meeting points’ (Argon, 1957).
We shall discuss temporary and permanent silk lines
later, but it is important to note here that the bridge
thread and the frame are frequently reinforced per-
manent parts of the orb web. They continuously
demarcate and preserve the perimeters of the site,
whether or not an orb is present. The frame silk may
be relatively thick, and it represents a substantial
energy expenditure, but the bridge and frame are
long term investments, supporting the hypothesis that
once sites are established they seldom change
(Peakall, 1971).
Building-Time
Like humans, spiders build both temporary and per-
manent structures. While the silken material, as far as
it has been analyzed, is similar in both web categories,
the design in each shows distinct differences.
Economy of labor and material is the outstanding
characteristic of the short term, daily-renewed web,
while more time, material and effort are expended on
a structure which serves for weeks or months.
Another interesting feature of the two design types is
their functional separation: the temporary structure
serves as a highly efficient trap which catches prey
for one individual, while a web with a permanent
design frequently serves several spiders for many
functions, such as feeding, protection, or raising
offspring.
The best representative of a short term building is
the geometric orb-web of Araneus diadematus. It
shows structural simplification through symmetry,
which Nervi (1956) postulates for economy of mater-
ial in building, and which is essential in a frequently
renewed structure. It has been shown (Witt, 1952)
that laying spiral sections perpendicular to the next
radius, which results in a logarithmic spiral, consti-
tutes the shortest and simplest route for the builder. If
we think only of economy in material, the logarithmic
spiral is wasteful, because a good device for catching
and holding air-borne prey should have narrowly
arranged spiral turns at the periphery where support-
ing radii are far apart; and a narrow spiral is
really quite unnecessary near the hub. Evidently
economy in movement and orientation are more
important for the spider in a temporary structure
than design features concerned with prey capture
alone.
Each day Araneus diadematus constructs a large
aerial net for flying insects. In comparison Cyrtophora
web/spider complexes, which claim a smaller surface
area, are built on a permanent basis. These are not
merely chance variations in building, they represent
two major strategies of architectural design and sup-
port. Norberg-Schulz (1945) concisely stated Tn
principle, we may distinguish between two types of
skeleton structures: embracing and repetitious. The
embracing skeletons are used to span large continu-
ous distances and mostly form a closed whole’ (Fig. 7,
and the Buckminster Fuller geodesic domes) (see
Interdisciplinary Science Reviews 1, 39 (1976)). The
orb uses an embracing scheme: while spokes can be
added to the cartwheel structure, the design cannot
be changed without upsetting its symmetry. Repetiti-
ous skeletons, on the other hand, can be enlarged by
adding units at the periphery without destroying
structural integrity. This design allows flexibility and
repeated additions, as represented in space and sheet
webs.
So the orb web, in contrast to space and sheet webs,
shows modules which consist of short sequences of
thread-laying behavior which are repeated several
hundred times whenever an orb is rebuilt. It is a good
illustration for Marcel Breuer’s words, that ‘the
search for simplification is, of course, connected with
a view to finding a prototype for mass production’.
The geometric orb, which covers the widest possible
INTERDISCIPLINARY SCIENCE REVIEWS, VOL 1, NO. 4, 1976 329
Figure 7. in building the exhibition hall in
Turin, Pierre Luigi Nervi used a modular
design, where precast structural elements are
employed many times, for reasons of
economy in material and labor. Though mesh
size varies in the vertical geometric orb web,
we can look at it as covering as large an area as
possible with as little material as possible;
construction is simplified by a behaviour mod-
ule, which is coded in the central body of the
spider’s central nervous system.
area with the least material and effort, owes its har-
mony to ‘the repetition of a module in such a way that
all parts of the structure coexist in simple numerical
relations’ (Vitruvius, 1940).
We can imagine the evolution of the orb web as a
refinement of the modular design; that is, as a short
sequence of probing and thread positioning move-
ments which can be repeated at rapid speed many
times. As a matter of fact, the spiral-laying Araneus
diadematus usually forms more than 1000 meshes in
rapid sequence in less than 20 minutes, occurring as
repeated execution of a relatively simple pattern of
behavior which is programmed in the spider’s central
nervous system. Destruction experiments (Witt,
1969) have shown that the central body at the rostràl-
caudal end of the supraesophageal ganglion of
Araneus diadematus may be the site of the modular
code. Laser lesions here result in severely disturbed
web regularity.
The exhibition hall in Turin is a human example of
modular design. Tn the building of the Exhibition
Hall in Turin (1948-49) Nervi employed precast units
of reinforced concrete for the barrel-shaped vault of
the hall, their maximum thickness being scarcely two
inches. With these thin sectioned components he
bridges an area 80 m long, an astonishing ratio
between expenditure of material and performance.
The corrugated arrangement of these prefabricated
units forming the ribs of the roof gives them the
necessary strength and at the same time solves most
handsomely the problem of direct lighting’ (Joedicke,
1959) (Fig. 7). For this structure, as well as for an orb
web, the equivalent of a temporary scaffold is neces-
sary.
Prospective Duration
Peakall (1971) describes another measure which
Araneus uses to conserve energy and material in the
frequently renewed orb-web. He placed ‘cold’ spiders
on radioactive webs, which they readily accepted.
After using the foreign webs for the rest of the day,
they carefully took them down piece by piece and
ingested the old silk, as this species usually does. In
webs built subsequently the reappearance of the
amino acids, the building stones of the old silk, could
be quantitated by measuring radio-activity. In every
case more than 90% of the old silk radioactivity was
present in the web two days thereafter, illustrating the
common use of recycled building material, and
reminiscent of repeated employment of components
of short term human structures.
When a backpacker buys a tent as a short term
shelter, he selects a system with a short set-up time,
and the equipment which permits quickest erection is
frequently chosen over a more lasting model. In a
similar way, Araneus lays its threads with enormous
speed during the daily web construction, fusing one
thread to another at about 2000 points in 20-40
minutes. For the human observer orb web building
looks as if a well prepared plan is carried out in a
systematic fashion, while space web construction fol-
lows a predominant trial and error procedure. In
sheet and space webs, there does not seem to be a
precoded neural template; both Cyrtophora and Mal-
los take several days to build their first web, then
continue adding parts.
Cyrtophora changes between rapid positioning of a
thread, extensive probing, and testing, and frequently
separates an already positioned strand and fastens it a
second or third time at a different place (Wiehle,
1928). Observers of Cyrtophora web building
describe how old silk is discarded whenever repair or
renewal takes place, rather than the web being
reused. Thus,Adn the space and sheet web, short
building time and reingestion of material are aban-
doned in favor of a more permanent structure. Even
after a rainstorm sheet and space webs are still used,
whereas orb webs are destroyed.
In the repetitious type, structures can be more
flexible, and Metepeira and Cyrtophora show a wide
variety of forms. Because of multi purpose flexibility,
these systems can accomplish a diverse array of func-
tions not available to the orb web. For one thing the
spider can move in three dimensions while, for
another, the tangle provides a degree of protection
against predators for both spider and eggs. In addi-
tion the newly hatched young ‘exercise’ on the upper
threads. Most importantly, the site is preserved on a
permanent basis. Although one Cyrtophora may
exchange webs with a çonspecific, other spiders can-
not colonize the now-occupied space.
We might consider the permanent strategy as a
homestead. Once a squatter develops his property he
forces others off and exercises control over its use. In
this sense he has the advantage of a home base free
from outside interference. If he then builds a basic
home, like a log cabin, the settler will be able to
occupy the simple structure and have the option of
adding extra rooms as they are needed. This is one
reason why many large American homes still have at
their center a single room cabin or other primitive
form of dwelling.
BEHAVIOR AND DESIGN
In our four spider species and their web complexes
there is almost a continuum in the degree of social
interaction. The orb web of Araneus diadematus
appears to be designed for single occupancy; there is
only one small area, the hub, from which the web can
be monitored; the converging radii direct all vibra-
tions and locomotor pathways to this single spot.
Little inter member tolerance exists in this species.
Whatever produces vibration in the web is attacked»
be it a fly, a sibling, or a tuning fork.
Even solitary animals, however, have some contact
with their species—minimally with mates and young.
The web of Araneus diadematus is the substrate for
courtship, initiated by the wandering males who
pluck or strum with a leg on the periphery of the
female’s web. Before actual mating takes place the
male connects its own thread to the web of the female*
I and so before two animals get together they must add
another structural element. When hundreds of young
hatch from their egg case they stay together for some
time, not on an orb, but on a communally constructed
sheet (McCook, 1889; Burch, in preparation). In
every case the orb functions for a single spider, and
the group interactions are relegated to another struc-
ture.
In contrast, Cyrtophora and Metepeira exhibit reg-
ular conspecific interactions, which take place on the
usual web structures; additional features are not
built. In Cyrtophora, if webs are found touching,
neighbors may steal food or exchange webs, and
predation on each other is possible but rare. When an
egg sac hatches, spiderlings proceed directly to over-
hanging space webs. They build their own structures
in the mother’s web and animals within the complex
remain tolerant for several days. They probably
aggregate around webbing, rather than around each
other. Courtship, however, is dependent on the pres-
ence of the female and not her web.
Like Cyrtophora, Metepeira spinipes are always
found in groups, exhibiting characteristic spacing
(Blanke, 1972; unpublished measurements by J.
Wesley Burgess). The space web in Metepeira is the
arena for social interactions. Webless males may wait
h here to steal food from neighbors’ orbs, or descend to
retreats for courtship and mating. Here the young go
after hatching, and the space web also serves for the
attachment of their first webs. Males may cohabit for
days with unmated females, in the area above the
retreat. Curiously, after mating, the egg sac is built
above the retreat and this area is filled.
Because Mallos enjoys a wide spectrum of social
interactions, the web forms a permanent substrate.
As a product of communal spinning, the sheet allows
aggregation, group predation, and the joint rearing of
young. Tolerance is complete. Males approach
females in a simple courtship and tiny immatures run
on the web, feeding unmolested on prey caught by
adults. It is not known yet how males and females
select each other on the communal structure, where
all animals appear in close contact at any time, but
there may well be some undiscovered structural fea-
tures which serve mate selection.
In both the sheet and space web the repetitive
construction system is used, and there is an increase in
web size for additional colony members. Similarly,
instead of building a separate nursery for a family, an
architect may prefer to provide regular adult-sized
rooms with juvenile furnishing, knowing these rooms
will serve many ages in the years to come.
Another way to look at social behavior is to find out
how activities in a web/ spider complex are organised.
A single Araneus diadematus, conserving reusable
structural material, may coordinate activities herself.
Thus, the catching web, egg sac and retreat area, are
well separated. Likewise Cyrtophora males are not
dependent on web structure to mate but use cues
directly from the female.
If the site regularly includes several members, how-
ever, it may be more efficient to coordinate group
activities with something structural, such as the web
itself. We might say that at home we know our way
around very weffl but in the expanse of a big city an
individual’s need for maps, direction signs and well
marked roads becomes important.
The behavior of Metepeira is largely regulated by
its web structure. Prey catching on the orb resembles
Araneus diadematus, but distance between orbs is
fixed by a permanent space web. Orientation of males
to females, rather than taking place in the open, is
directly channeled through the overhead roadway of
the space web, which connects site members. As in
Cyrtophora, dispersal of young Metepeira is directed
by the space web, but the young stay around for a long
time and may disperse over the entire colony-shared
web. The locus of every spider action is thus pre-
served within the complex: the temporary orb is fixed
to the permanent space web, while the retreat pro-
vides a resting point for the male and also preserves
inner space for the egg sac. In a changing environ-
ment the web complex is its own urban zoning system,
under a plan which was developed over time and is
encoded in the spider’s genes.
The Mallos plan is less flexible, turning most func-
tions over to structural integration. On a shared web
substrate site members travel on silk pathways laid
down earlier, as the draglines of other walking colony
members. Not only movements, but also communal
predation signals are carried by the surface sheet,
whose threads exhibit a resonance response, which
INTERDISCIPLINARY SCIENCE REVIEWS, VOL 1, NO. 4, 1976 331
mediates the predatory cue (Burgess, 1975). Within
the web, chambers for egg sacs and pregnant females
centralize some aspects of reproduction between
many members. Because the sheet is asymmetric and
continuous, groups of spiders can catch prey together.
Flies are possibly attracted to the web (Diguet, 1915).
STRUCTURES OF COMMUNICATION
Animal communication is easy to talk about and hard
to study, mainly because our everyday exchanges
involve questions like ‘who talks to whom?’ or ‘did I
get through?’ To investigate communication scien-
tifically, we need something we can measure.
Behavioral scientists may look at information trans-
fer, which consists of any measurable communication
units which are directed into the environment by an ,
organism. In looking at information transfer, we
implicate structure and design as substrate for trans-
fer of communication units. Also, since signal energy
deteriorates over distance, animals must arrange to
be in resolving range. To widen this range they pro-
duce some signal transport network. Like a spider in
her web, every human is surrounded by structural
extensions of his senses; we are interconnected by
telephone, and telex wires, radio waves, roadways,
railways, postal ways and air ways. We call our neigh-
bor up, send a letter, exchange books, or walk
through halls and over roads to see him personally.
We are directly tied to our conspecifics by the struc-
tural networks of our civilization.
Spacing is one social factor dependent on com-
munication. Unless they distribute completely at ran-
dom, animals either aggregate or spread apart, and
these distribution patterns are dependent on infor-
mation exchange between individuals. In this sense
many apparently solitary animals communicate with
their conspecifics, giving signals necessary to preserve
distances. There is likelihood that this is so in the
single orb web builders.
In order to eat, spiders must receive some informa-
tion from their potential prey, usually in the form of
thread vibration. Here the orb structure organizes the
environmental information by filtering out only prey
of appropriate size (Witt, 1975), and dampening low
vibrations or aerial sounds (Fink, et ai, 1975; Szlep,
1964; Walcott, 1969). In a social context, the reso-
nance of the Mallos web provides for communal
predation, because the web carries a clear signal when
a trapped fly buzzes (the communal predatory cue),
but dampens the vibrations of walking spiders
(Burgess, 1975).
Wherever web connections are shared between
animals, some vibratory signals will be transmitted.
Thus, while two touching Araneus diadematus orbs
frequently result in cannibalism, orbs of the colonial
Metepeira are separated by a tangle of space web.
These functions have many counterparts in the
societies of other living beings. As a structure builder,
man designs houses to connect the units of his family,
but includes doors and walls to contain individuals
and to provide needed privacy. A company building
may be designed to channel people to the executive’s
office, but inevitably there is a secretary present, who
coordinates and filters the flow of traffic. Streets with
smooth surfaces connect human habitations and per-
mit circulation in vehicles or with soled shoes. Silk
lines connect spiders for movement on hooked feet.
FACTORS INFLUENCING DESIGN
Design in human buildings and spider webs reflects
the immediate functional requirements as well as
traditions. Houses in suburban developments must
provide the inhabitants with shelter of the proper size
and with a comfortable environment, but they also
contain non-functional elements like white columns,
ornamental trims and gables, which are reminiscent
of Greek temples. Similarly, features in the design of
spider webs are explainable either through ‘tradition’
or through the immediate advantage they offer the
builder.
We are not sure that the term ‘tradition’, which
Webster defines as ‘designed with conscious adher-
ence to architectural styles of the past’, strictly applies
to spiders. Another name sometimes used is ‘genetic
inertia’. Their central nervous system, which signals
to the legs and the rest of the body to move in a
specific way so that silk is laid to form the web pattern,
develops according to a genetic code. This code is
passed on from generation to generation and,
although subject to mutation and natural selection in
individuals, for the species overall it is largely
unchanged for long periods of time,
i I^Jfo explore the extent of previous coding, investiga-
tions have been made to determine whether spiders
reflect individual experience in the web pattern: fol-
lowing earlier efforts, Reed, et al. (1970) measured
web patterns of growing Arnaeus diadematus litter-
mates, half of which had caught flies in their web
daily, while the other half had been exclusively fed by
mouth. One could speculate that non-use of the orb
web as trap for prey would result in decreased atten-
tion to detail in construction, perhaps giving rise to
less regular, wider-meshed webs. However, no differ-
ence could be found between the webs of fly-catching
and those of mouth-fed spiders, indicating the mini-
mal influence of experience on fine structural design.
Other experiments with spiders raised under different
conditions confirm this conclusion. One can infer that
non-use of structural elements in spiders will lead to
their disappearance even more slowly than in human
building.
On the other hand, we have made pictures to show
that some novel environmental contingencies are
reflected in design (Fig. 3). These photographs were
made when spiders were confined to cages in which
they could not select an appropriate site; such a
332 INTERDISCIPLINARY SCIENCE REVIEWS, VOL. 1, NO. 4, 1976
condition may never occur in nature, where they can
change web site according to available open space.
Under these somewhat artificial building conditions,
mechanisms exist which adjust to immediate require-
ments in web-building spiders.
Partially in an effort to test the effects of extreme
environmental changes on web pattern, and partially
to find out whether behavior of an invertebrate ani-
mal can adjust to weightless conditions which have
never been experienced by the species before, web-
building spiders were sent into space to Skylab II as it
circled the earth. Two animals were carried to the lab
in small vials in the astronauts’ pockets; they were
released into cages, and were monitored photo-
graphically. Four days after release from the vials the
first animal started to build a web which was about as
large and the silk as regularly spaced as if it had been
made on Earth.
The webs were distinctly different from earth webs
only in radial angle distribution, thickness of thread
and number of turning points in the spiral, all indi-
cators of orientation to gravity (Witt, et al., 1976).
One can compare the activities the spiders and
astronauts performed under weightless conditions:
the latter went through extensive training, read and
thought about how to adjust to the expected new
conditions, while the spiders had to cope with every-
thing completely unprepared. Both solved the prob-
lem of moving around and structuring their environ-
ment in the usual way, compensating efficiently for
the absence of gravity. This indicates that comparable
results can be achieved by the two different organ-
isms, each approaching the problem in his own way.
There are no fossil webs preserved, so we do not
know how the selection process has affected web
design in phylogenetic spider history. The surprising
observation is that only a few basic structural designs,
like the orb, the sheet and the space pattern, are
repeated time and again in various combinations by
thousands of web-building spider species. It has given
rise to speculation that they are so good and unique a
solution to the functional problem, that they have
been discovered several times independently by
different species (see Kullmann, 1972).
Another way to explain the many appearances of
few designs in various species would be to assume the
existence of an ancestral web-builder which was a
subsocial animal with a composite web complex, simi-
lar to that now found in Metepeira. With development
through the millennia, increasing specialization
occurred: some species developed into single hunters
with very large, regularly and quickly constructed
orbs, others into social beings where communal
asymmetric continuous webs satisfied their needs
best, and others again into semisocial animals with
mixed web structures. Robinson and Robinson
[ (1975) call such a phenomenon ‘web development
with progressive reduction in complexity’. In contrast
to men, who preserved the flexibility to live alone in
family or communal groups, housed in single or
multifamily dwellings, present spiders are largely
born into their social and web patterns, which are
species-specific, coded in their genes.
Pattern changes throughout the lifetime of an ani-
mal are another indirect piece of evidence that the
various designs of spider webs have a common origin.
Newly hatched Araneus diadematus spiders con-
struct and live communally on a space web. As long as
they stay on this structure, they show mutual toler-
ance, even though they already have the ability to
catch and wrap small prey. After one to two weeks,
single animals will leave and build a first, perfect orb
web of their own; on this new web all signs of toler-
ance disappear, and littermates are attacked and
killed if they stray onto the orb.
One has to conclude that here the ability to con-
struct space webs and geometric orbs are both geneti-
cally transmitted to every Araneus diadematus indi-
vidual, becoming manifest at different periods in their
lives. If the animals build two different types of web in
their lifetimes, this implies a change in the web-
building ‘program’ contained in the central nervous
system. It is important to remember at this point that
many other factors, such as body shape, are consi-
dered when a phylogenetic family tree of spiders is
constructed: our interest at present is only to look at
the web design as one component for identification of
relationships.
The choice of material influences design. As steel
beams or prefabricated units of reinforced concrete
become available, new possibilities are opened in the
construction of building spaces. Spiders, in contrast,
have apparently kept to one material in all their
structures, a polypeptide: silk. Silks of very similar
composition have been used widely, even by animals
as remotely related to spiders as insects, mainly moths
and butterflies, the most widely known being the
larvae of the moth Bombyx mori, the silk worm. Lucas
and Rudall (1968), reviewing studies of the silk of the
orb-weaving spiders Argiopidae, have compared silks
from various glands in the same species with silks
produced by other species.
The thread which bears the weight of the spider and
maintains the tension of the web contains a high
percentage of short side-chain amino acids: Peakall
(Witt et al., 1968) gives for alanine figures between
32.7 and 33.4g/100g silk, for glycine 24.3 g/100g
silk, and serine 6.3 to 6.4 g/100 g silk. 10 or 11 other
amino acids with longer side-chains make up the rest
of the silk. A comparison shows that the strength of
dragline silk with 7.8 g/denier tenacity is almost as
high as that of nylon with 8.7 g/denier: but at the
same time the extensibility of spider silk is considera-
bly higher. To accomplish similar ends, both spiders
and humans have developed comparable processes.
With the building of the Brooklyn Bridge, a suspen-
sion bridge completed in 1883, the Roeblings, father
and son, developed a device for spinning steel strands
on the job into great cables and used these in crossing
vast distances with majestic grace. Steel thus was used
in tension, consistent with its inherent nature’ (Lloyd
Wright, 1962).
INTERDISCIPLINARY SCIENCE REVIEWS, VOL. 1, NO. 4, 1976 333
Comparing drag-line silk, which has to bear the
weight of the animal, with cocoon silk, which has
more protective-isolating functions, Lucas and
Rudall (1968) conclude that the mechanical prop-
erties of these two silks made by the same animal are
appropriate for their function in that the drag-line has
a very high tensile strength, while that of the cocoon is
only moderate. After developing such optimal mater-
ials, genetic inertia insured that spiders would use
them for many generations. Or looking at it in
another way, the development of strong and elastic
silk has made it possible to produce the current web
designs; but the inherited pattern of silk synthesis has
thereafter restricted the possible variety of web struc-
tures.
Under human conditions, material restraints can
be observed in Greek temples. The stone structures
followed post and beam construction, which was
developed with the older material, wood. Spaces
became limited by the length of stone beams, and
columns had to be spaced close together, resulting in
massive buildings. Only generations later were t«nie
arch and vault developed, making it possible to build
greater spans out of stones; and the light G«3|mc
cathedrals could be constructed by using the ‘new’
material in an adequate way. Stone was eventually
superceded by steel and reinforced concrete, which
again led to new developments numan building
design.
CONCLUSIONS
We have analyzed a small sample of spider webs!
deliberately selected for Ils grea®arieH. We have
tried to explain the characteristic design oSeaH
structure by the functional requirements. Parallel!
were drawn between the design of human andi)ider
structures, and in many instances it was possible to
compare structural details with each other and find
similar underlying engineering principles.
Like humans, spiders build temporary as well as
permanent structures, the former requiring a plan
which can be executed quickly, the latter being built
more slowly, with options for later additions. The
embracing modular design has been found economi-
cal in both living beings for certain building require-
ments; the repetitious design for others.
Individuals have to communicate and require a
certain amount of privacy. Both needs can be traced
in the buildings constructed by spiders and by men.
There are walls to separate individuals, as well as
pathways for access to each other; and communica-
tion over distances is achieved in both kinds of struc- I
tures through specialized elements, like vibrating silk
lines or electric wires.
Designs reflect clearly the degree of social behavior
which the inhabitants exhibit. Single living beings are
housed differently from families and fraternal groups.
Offspring can be raised in the parents’ structures, or
can occupy their own shelter, designed to serve only
the babies’ functions.
Spiders’ space, orb, and sheet webs have been
shown to be distinctly different in design as well as
function. They represent different adaptive
strategies, and have their parallels in human con-
struction. We have seen that they can occur by them-
selves, or in combination. Thus many functions can be
served I® one building through the integration of
designs.
Even the obvious differences between arachnid
and human structures, like building material and
coding of design, can be seen as showing some
similarities m underlying system principles. The
natural selection process for the most efficient spider
web is comparable to economical considerations
which enter an architect’s mind when he designs a
building. Materials, though more uniform in spider
structures than^H human buildings, have been j
selected in both examples for efficiency, and have ”
consequently dictated structural layouts.
One of the most interesting features of spider
webs is theiBnteraction with the environment. The
surroundings influence web structures, but, in return,
web structures alter the area in which they occur.We
have only just started to investigate such questions.
The understanding of this interaction between build-
ing, ÉÉmder, and the environment may be the area in
which we human habitat builders can learn most in
the future from observing spider web designs more
closely.
Acknowledgements
The authors thank R. Daniels, M. Scarboro, E. H. Williams,
C. F. Reed, R. Jackson and the National Science Founda-
tion for support and assistance.
LITERATURE CITED
Where a recent review was available, we quote this rather
than all relevant papers ever published on the subject matter.
G. C. Argon, Marcel Breuer: disegno industrialo e architect-
tura; Gorlich, Milan (1957).
R. Baltzer, Beitraegezur SinnesphysiologiederWebespinnen,
Mitt. Naturf. Ges. Bern. 163-187 (1923).
R. Blanke, Untersuchungen zur Oekophysiologie und
Oekethologie von Cyrtophora citricola Forskal in
Anadalusien, Forma et Functio 5,125-206 (1972).
J. W. Burgess, The sheet web as a transducer, modifying
vibration signals in social spider colonies of Mallos gregalis,
Neuroscience Abstracts, New York (1975).
J. W. Burgess, Social spiders, Scientific American, 234,
100-106(1976).
L Diguet, Nouvelles observations sur le mosquéro ou nid
d’araignées sociales, Bull. Soc. Nat. Acclim. France. 240-249
(1915).
F. Enders, Vertical stratification in orb web spiders and a
consideration of other methods of coexistence, Ecology 55,
317-328 (1974).
A. Finck, G. M. Stewart and C. F. Reed, The orb web as an
acoustic detector, J. Acoust. Soc. Am. 57, 753-754 (1975).
W. J. Gertsch, American spiders. Van Nostrand, New York
(1949).
M. Holzapfel, Die nicht-optische Orientierung der Trich-
terspinne Agelena labyrinthica (CL), Z vergi. Physiol. 20,
55-115(1933).
J. Joedicke, A History of Modern Architecture, Praeger, New
York (1959).
E. J. Kullmann, Beobachtungen des Netzbaues und Beitraege
zur Biologie von Cyrtophora citricola Forskal, Zool. Jb. Syst.
86, 181-216(1958).
E. J. Kullmann, The convergent development of orb webs in
cribellate and ecribellate spiders. Am. Zool. 12, 395-405
(1972).
F. Lloyd Wright, Architecture, Man in Possession of his Earth,
Doubleday, New York (1962).
Y. S. Lubin, Web structure and function: the non-adhesive orb
web of Cyrtophora moluccensis (Doleschall), Forma et Func-
tion, 337-358(1973).
F. Lucas and K. M. Rudall, Extracellular fibrous proteins: the
silks, Compreh. Biochem. 26B, 475-558 (1968).
H. McCook, American Spiders and Their Spinning Work
(1889), Pub. by the author, Philadelphia (1903).
P. L. Nervi, Structures, Dodge, New York (1956).
C. Norberg-Schulz, Intentions in Architecture, MIT Press,
Cambridge, Mass (1945).
D. B. Peakall, Conservation of web proteins in the spider
Araneus diadematus, J. Exp. Zool 176, 257-264 (1971).
F. 0. Pickard-Cambrldge, Arachnids, Arancida, Biol. Centr.-
Amer. Zool 2, 425-464 (1903).
C. F. Reed, P. N. Witt, M. B. Scarboro and D. B. Peakall,
Experience and the orb web, Deveiopm. Psychobiol, 3,
251-265(1970).
M. H. Robinson and B. Robinson, Evolution beyond the orb
web: the web of the araneid spider Pasilobus sp„ its structure,
operation and construction, Zool. J. Linn. Soc. 56, 301-314
(1975).
R. Szlep, Change in the response of spiders to repeated
vibrations. Behaviour 23, 203-239 (1964).
A. L. Turnbull, Ecology of the true spiders, Ann. Rev. Entomol.
18, 305-348 (1973).
M. Vitruvius, De Architecture, London (cit. from Norberg-
Schulz) (1940).
C. Walcott, A spider’s vibration receptor: its anatomy and
physiology. Am. Zool. 9, 133-144 (1969).
H. Wiehle, Beitraege zur Biologie der Araneen, insbesondere
zur Kenntnis des Radnetzbaues, Zschr. Morph. Oekol Tiere
11, 115-151 (1928).
P. N. Witt, Ein einfaches Prinzip zur Deutung einiger Propor-
tionen im Spinnennetz, Behaviour 4,172-189 (1952).
P. N. Witt, Behavioral consequences of laser lesions in the
central nervous system of Araneus diadematus Cl, Am. Zool.
9, 121-131 (1969).
P. N. Witt, The web as a means of communication, Biosci.
Comm. 1, 7-23 (1975).
P. N. Witt, C. F. Reed and D. B. Peakall, A Spider’s Web.
Problems in Regulatory Biology, Springer, Berlin (1968).
P. N. Witt, M. B. Scarboro, R. Daniels, D. B. Peakall and R. L.
Gause, Spider web building in outer space: Evaluation of
records from the Skylab spider experiment, J. Arachnol. in
press (1976).
The manuscript was received 8 March 1976.
B) 1976. Heyden and Son Ltd.
INTERDISCIPLINARY SCIENCE REVIEWS, VOL 1, NO. 4, 1976 335