Images Collection



View this article in Search Friendly Plain Text
NOTE: This plain text article interpretation has been digitally created by OCR software to estimate the article text, to help both users and search engines find relevant article content. To read the actual article text, view or download the PDF above.
Biosciences Communications
Editor: Stacey B. Day, New York, N.Y.
Publishers: S. Karger AG, Basel
Separatum (Printed in Switzerland)
Biosci. Commun. 1: 7-23 (1975)
The Web as a Means of Communication1
Peter N. Witt
Department of Human Resources, Division of Mental Health Services, Research
Section, Raleigh, N.C.
Key Words. Spider • Orb web – Web architecture • Silk • Signals • Receptors * Lyriform
organs • Chemosensitivity • Mating – Early development
Abstract. The radial structure of the orb web with the spider in the center, and the
tightly stretched polypeptide material with ifs:specific chemical and physical properties, are
discussed as efficient transmitters of various signals to a number oMpecialized sense organs
on thé spider’s body. An animal’s behavior in response to a signal pi dependent on the nature
of the signal and on age, sex and species of the receiver. An overview of the research
literature endeavors to separate a small amount of evidence from a large array of speculation
and surmise, in regard to the orb web as a means of communication, mainly in the cross-
spider, Araneus diadematus Cl.
Introduction
Communication has been defined as the act of imparting or transmitting
information, the facts or information communicated, or the access between
persons or places (Websterm965). This paper is limited to discussion of the orb
web as a means of communication in spiders. If one observes the spider turning
toward a fly which has just struck the web, in contrast to the same spider sitting
unresponsive on the surface of a table while a fly walks by (Baltzer, 1923) — or
if one observes the elaborate approach and retreat ritual elicited by the male
spider’s drumming on the female’s web — one can hardly have doubts about the
1 Support to the author’s laboratory from the National Science Foundation and help
in the preparation of this manuscript by Mrs. Rubenia Daniels; Mrs. Mabel B. Scarboro and
Dr. Charles F. Reed are gratefully acknowledged.
Witt
8
important role which the web plays in imparting or transmitting information; it
constitutes an essential route of access to the spider’s perceptual systems.
There is much less certainty about the nature of the signal. It may be asked
whether the web transmits and the animal receives just one signal which puts it
on alert, or whether there are specific frequencies which are recognized as signals
of danger, and others as challenges for attack. Do the eight legs lying on dif-
ferent threads provide the means for establishing the direction and distance of
the prey? And how much is actually transmitted by the web, how much by
other means such as airborne sound? As one looks at the results of experiments
and observations, one finds an abundance of predominantly circumstantial
evidence, much of which is in my judgment quite convincing for the designation
of the web as a means of communication. However, there is an absence of
reports on the investigation of the whole chain of events, starting with the origin
of the signal, its transmission, its reception, and final analysis by the receiver. If
the animal has an organ particularly sensitive to vibrations of the substrate, and
if the strands of the web will transmit the struggles of the prey, and the animal’s
behavior indicates that it reacts to the movements of the prey, one could assume
that vibrations transmitted through silk strands of the web are signals containing
information for the spider about prey. Although investigations may alter details,
it will be shown in the following, on the basis of circumstantial evidence only,
that it has to be assumed that the web forms a substantial means of communica-
tion between spider and spider, between spider and prey, and between spiders
and their environment2.
The Web as a Substrate for Communication
Web Architecture
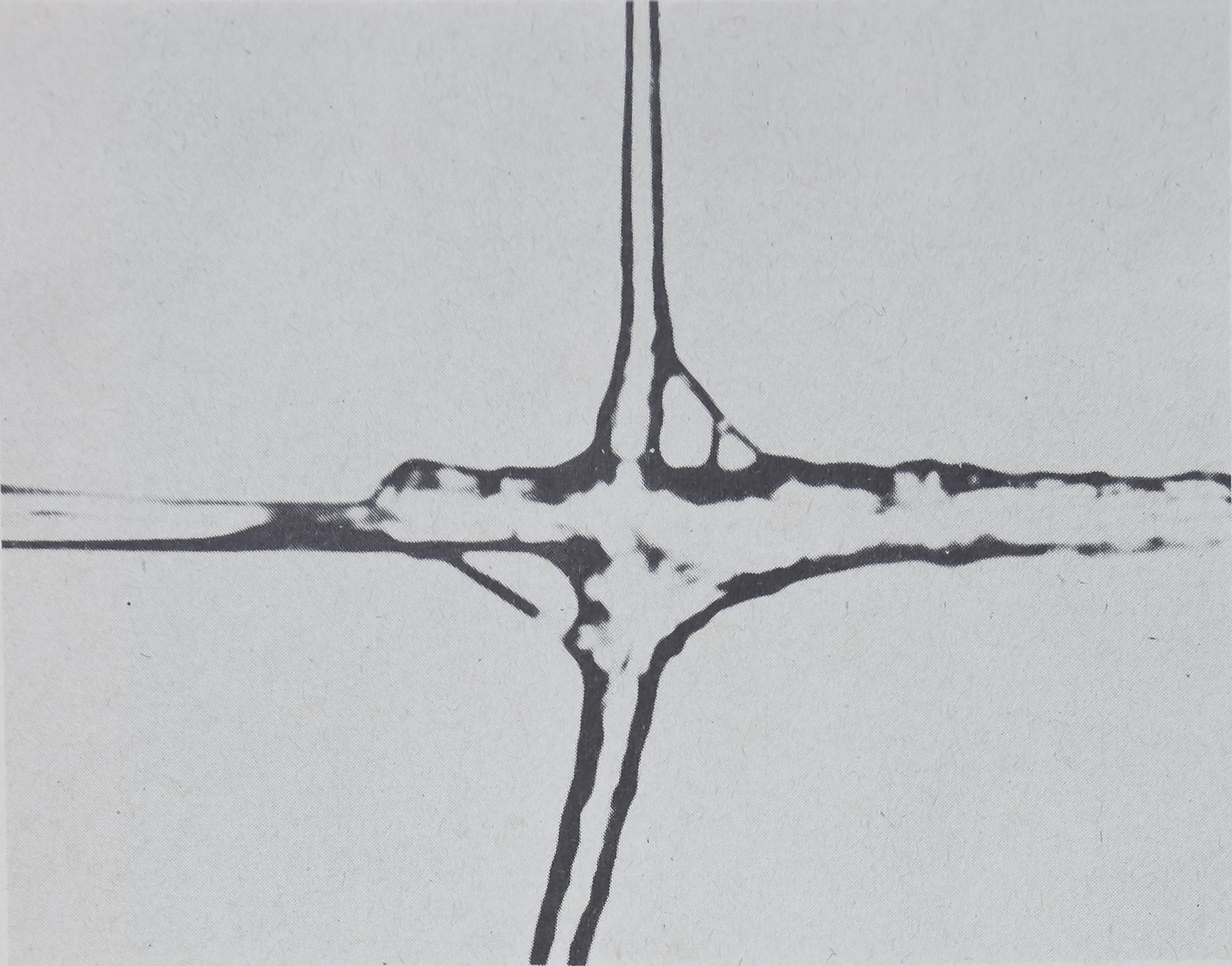
.A look at an orb web photograph (fig. 1) shows specific components of the
structute as most suitable candidates for reception and transmission of informa-
tion to the centrally located spider. All orb webs are variations on the same
theme, with a slightly larger or smaller central area, or with more or less oval
shape {Witt et al, 1972). The signal thread, which in some webs connects the
centrally located hub with the legs of a peripherally hidden spider (fig. 2),
appears to make the function of the radial structure even more obvious. The
radii function as extensions of the sensitive eight legs of the spider, and so for a
nearly blind animal enlarge the perceptual space. The communication advantages
2 Most observations have been made on Araneus diadematus; where not specifically
mentioned, this species was observed.
The Web as a Means of Communication
9
Fig. 1. This web of an, adult female Araneus diadematus or cross spider is one of many
which the animal built in the laboratory in the early morning hours during a period of about
30 min. The three parallelBfcs of the scale in the upper corner indicate the direction of
gravity and 20-mm distance between each two of the three threads. Observe the radial
structure of the orb, with the spider in the hub, and the suspension of the frame in the
environment. ,
of radial construction can be appreciated if one contrasts it with the inefficiency
of signal transmission to one point in a web which consisted of a rectangular
grid, in which the threads formed small squares.
If we touch a vibrating tuning fork to one radius of the web (Boys, 1880), a
centrally located spider will immediately turn in the direction of the fork. It will
run out toward the fork, and it may evefi wrap some swathing silk around its
prongs. If less hungry, the animal may apply only short pulls to the radius while
it stays in the hub. It has been shown (Peters, 1931, 1933; Bays, 1962; Robin-
son, 1969), that a weighty object like a glass bead, when properly vibrated, bears
sufficient resemblance to a struggling prey to bring a hesitant spider out to the
object. The radii put the spider virtually ‘in touch’ with the fly.
Such observations leave hardly a doubt that the web architecture plays a
role in communication between prey and predator and, as will be seen later,
between mature male and female of the same species.
Witt
10
Fig. 2., Another species of spider, Zygiella-x-notata, leaves one sector of the orb web
free of spiral threads. A signal thread connects the hub with the retreat. During most of the
day the spider sits in the retreat, front legs on the signal thread, and waits for prey to get
entangled in the web. Vibrations are presumably transmitted from any part of the web
through the radii to the hub and from there through the signal thread to the spider’s legs.
Other parts of the orb web, i.e., the frame and the sticky spiral, probably
play a supportive or at least a non-inhibitory role in communication. The frame
suspends the whole cartwheel pattern under balanced tension in the air and quite
effectively buffers against shifts in thç supporting structures. Observations of
frame construction show an animal’s efforts to revise the structure repeatedly
through shortening of frame threads (Salzmann and Witt, 1973). An Argiope
aurantia web, anchored only in grass, survives a heavy breeze showing that frame
threads are good mediators between a shifting environmental structure and a
relatively rigidly, fine-meshed net.
The sticky or catching spiral, which crosses the radii at angles close to 90°,
is thinner and more elastic than the radial thread, and provides minimal inhibi-
tion to the transmission of vibratory signals along the radii. That the thinness of
a strand is important is stressed by Jackson”s (1971) close-up photographs of
radius-spiral crossings: a thickened sleeve gives strength to the connection and
adds only a little rigidity to the transmitting radial threads (fig. 3).
The Web as a Means of Communication
11
Fig. 3. In this close-up photograph by Jackson (1971) the radius runs from right to
left, the spiral vertical, the periphery of the web is toward the right. Note how a relatively
thick sleeve of protein material reinforces the crossing of the threads, while the silk in
between is thin and has an even surface.
The spider’s practice of tightening the web, after prey has been cut out,
points to the importance of maintaining an even tension. It preserves the struc-
ture’s function as a transmitter; for prey-catching alone, a loose web would be
just, as effective as a tight one. Often no effort is made to close the hole which
results from detaching the prey; but as figure 4 shows, the periphery of the hole
is reinforced so that sagging of the residual structure is prevented. This periph-
eral reinforcing is only done for small repairs; more extensive destruction results
in partial replacement of the missing web part.
The idea had been proposed ([Finch, personal communication) that the web
may provide an early warning system for the approach of flying prey. It could
hypothetically amplify the air disturbances produced, by a slowly approaching
fly, and transmit them to the waiting spider. As far as I know, no such function
has up to this time been established (Walcott, 1969; Finch, 1972), and the
existence of a variety of hairs on the spider’s legs which are highly sensitive to air
currents makes such a function for the web unnecessary. If a vibrating tuning
fork is moved through the air toward a spider, it raises the front legs in the
direction of the fork as if probing for air-transmitted waves, rather than receiving
12
Mm
Fig. 4. The same web as in figure 1 was photographed after the spider had caught one
house-fly. Note the big hole in the net and the reinforcements which the spider has applied
to the sides of the hole: the undamaged part of the web is as tight and functional as before.
vibrations from the web. At our present state of knowledge there is no good
reason for assuming that the web acts as a receptor for airborne waves, and that
it provides an early warning system for the spider.
Some Relevant Information on Silk Chemistry and Physics
Only one type of gland, the ampullate gland, produces the dry silk for hub,
radii, and frame of the orb web ( Peakall,1964, 1968); other glands provide the
cover and/or the core of the viscid spiral thread, others the silk for the egg sac,
and others again the attachment discs and swathing bands. The product of the
ampullate glands should provide thread suitable for transmitting signals as well as
permit the spiders to run along the radii to any part of the web.
For amino acid analysis, Peakall either rolled out ampullate silk onto a
machine-turned glass rod and dissolved the product, or he collected web silk just
before the sticky spiral had been started. He was able to establish the amino acid
composition of structural silk, confirming earlier analyses of Lucas and Shaw
(1961). A predominance of amino acids with small side-chains (glycine, alanine,
serine) was found, with variations in ratio according to species. Such composi-
The Web as a Means of Communication
13
tion appears to be dictated by structural requirements of the silk; it does not
seem to reflect phylogenetic relationships of the builders. It can be deduced
from these data that the function of the structural silk strands is of prime
importance for the survival of the animal. One such important function is com-
munication.
Somewhere between the lumen of the ampullate gland and the emergence
from the spinneret a transformation takes place: the fluid silk is made into an
insoluble and strong thread. The process is not completely understood. War-
wicker (I960, 1961) concludes from X-ray diffraction photographs that his data
are best fitted by ‘random packing of antiparallel pleated sheets of polypeptide
chains’. The high tenacity of the structural silk (7.8 g/denier) and its consid-
erable elasticity (extension of 31 at break) {Lucas, 1964), can be explained as a
result of the macromolecular structure. If one considers the relatively large bulk
of an adult female Araneus diadematus spider with a weight of about 500 mg
dangling at the end of or pulling with full strength at a silk strand of 1—2 /i
diameter without breaking it, one gets an impression of the high strength and
elasticity required of the thread;
Of special interest are the calculations of Langer (1969) who finds that the
‘material properties of the silks are related to the structure and functions of the
spider webs’. He1 considers gravity and wind forces, but finds localized forces of
particular Interest. He treats mathematically two signal velocities on a silk
filament, a longitudinal velocity of about 2 km/sec and a transverse velocity
which is much slower. Langer concludes that a tug in one web element results in
a proportional response in every other element of the web, with a time delay of
the faster signal in the order of lOjusec. The separate circulation of the two
signals through the web to the spider’s leg could quickly provide information on
the nature, location, and sort of disturbance in the web.
While there can be hardly any doubt that’ the orb web qualifies as a good
transmitter of vibrations and tension changes, its part in providing chemical
signals has hardly been explored. Millot (1946) describes observations of Kaston
(1936) and experiments of his own which make it likely that a spider receives
chemical signals when it touches the web. According to Millot, a thread secreted
by a female excited the male of the same species on touch, even in the absence
of the female. If the silk was left in air for 2 days, or if it was dipped for 15 min
into ether, it no longer excited thp male. After evaporation of the ether, the glass
still elicited a reaction from the male. Millofs conclusion, that sexual behavior
of male spiders is released by a volatile chemical on the silk (and integument) of
the female, may not apply to all web-builders. It would be important to repeat
the experiment with several orb-web builders and carefully exclude all other
cues; increasing experience with pheromones {Blum and Brand, 1973) should
make it worthwhile to try to identify messenger substances. Organs which could
receive Millot’s chemical signals from the thread will be described below.
Witt
14
The Spider as a Receiver of Communication through the Web
Vision
It has been stated earlier that vibratory signals play the main role in infor-
mation gathering for a spider in the orb web. Several lines of evidence support
the view that vision is unimportant.
As far as can be established, the histological substrate for high visual activity
is lacking in web-builders. Hanstrom (1934) points out that web-building spiders,
in contrast to hunters, show a relatively poorly developed nerve mass for re-
ceiving and processing visual signals. On the other hand, web-builders, in contrast
to hunting spiders, have a relatively well developed central body in the central
nervous system, which apparently coordinates leg placement in web construc-
tion. Destruction of the central body by a laser beam was followed by a severe
distintegration of the pattern for all webs built afterwards (Witt, 1969).
Most spiders construct their webs in the dark and can use them for prey
catching during the night. In order to make sure that no residual light was
needed in web construction, the author compared 27 parameters in the geome-
try of orb webs built by spiders in the lighted laboratory with orb webs built by
the same spiders after they had been sealed into an opaque box: no differences
were found. Peters (1931) eliminated the eyes of web-builders and obtained
similar results: the orbs built thereafter appeared unchanged.
Lyriform Organs (Slit Sense Organs)
In a brief, comprehensive review, Barth (1973) discusses the structures of
these organs and deduces their function from structural principles. The receptors
consist of either one or several slits which are covered with a flexible skin in
which a nerve fiber terminates. The slits lie between rigid pieces of spider cuticle.
The skin is deformed as a result of pressure on the cuticle, and even small forces
applied to the cuticle provide deformation of the sensillum. Mechano-electrical
transduction is most likely caused by non-uniform compressional deformation of
the dendritic end of the receptor nerve, and the signal is analyzed in the central
nervous system.
An interesting feature is the distribution of the slit sense organs: over 100
have been counted, mainly along the eight legs of the spider, close to each joint
and near the pedicel. Seyfarth and Barth (1972) ascribe the kinesthetic orienta-
tion of their spider (Cupiennius salei Keys.) to these organs. They are able to
show, in selective destruction experiments, that spiders are impaired in their
ability to find their way back if specific slit sense organs are destroyed. The same
organs could play an important role in web construction and orientation in the
web. Consider the nearly blind animal (in the dark) apparently using information
on the position of all eight legs to probe the width of angles between threads and
thread distances. It is interesting to speculate that the input from more than
The Web as a Means of Communication
15
100 slit sense organs from all parts of the body could be integrated and analyzed
in the central nervous system in such a manner that the animal has an inner
‘picture’ of the position of all parts of its body at all times. Such function could
be compared to that of the human secondary visual centers, which integrate
information received from many points on the retina. It is difficult to explain in
any other way the number of slit sense organs and the well-oriented behavior of
the spider on the web.
The lyriform organs consist of a row of several slits. They are located on the
distal end of the metatarsus and appear to be sensitive vibration receptors
(Liesenfeld, 1961; Walcott, 1969). All investigators find a discrepancy between
the behavior of the animals and the electrophysiological records: while the prey
vibrates the web at relatively low frequencies, and while the spider reacts quickly
to frequencies well under 50 Hz, the electrophysiological reaction is definitely
most sensitive at frequencies oveft>100 Hz. It is even more puzzling when Bays
(1962) states that he could train his spiders to distinguish between the vibration
of two low frequency tuning forks, and Mark! (1973) finds^the electrical sensi-
tivity of spider-leg nerves nearly independent of frequencies below 100 Hz. One
may have to consider the complexity of the signal of varying amplitude and with
possible overtones which the fly elicits, together with input from other organs
which perceive, for example, airborne vibrations, to understand the behavioral
reaction of the spider to low frequency signals.
Trichobothria
Most hairs covering the legs of spiders lie closely along the surface; inter-
posed single bristles 100—700/i long stand at right angles to the cuticle. The
latter are called trichobothria; under a microscope it can be seen that they
vibrate slightly in the faintest breeze. Corner and Andrews (1969) have shown,
in non-orb web builders, that trichobothria react to airborne vibrations of low
frequency over short distances. Markl (1973) discusses the difficulties which are
encountered when one tries to distinguish between airborne and surface-
transmitted vibrations. For this discussion we can regard trichobothria as auxil-
iary perceptual organs for airborne waves which add to the information which
the spider received through the web.
Chemosensitive Hairs
It was considered above that the male spider responds to a possible chemical
transmitter on threads of the webs of mature females. Observation of prey-
handling by spiders leaves no doubt that the animals can distinguish between
bitter tasting (quinine), and sweet tasting (sugar), and many other solutions. But
until recently it remained an open question whether the spider had to swallow
the chemical in order to identify it.
W. Holden (unpublished film) took moving pictures in the author’s labora-
Witt
16
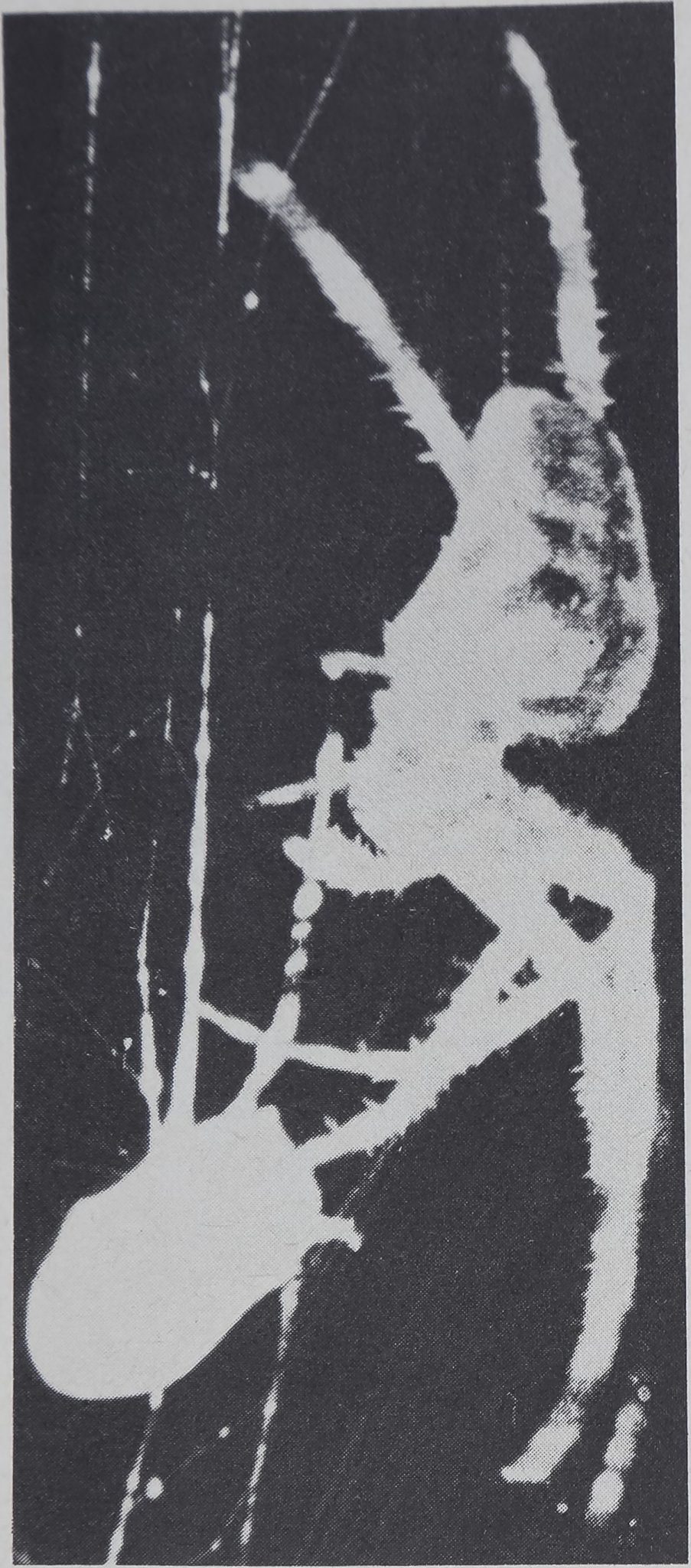
Fig. 5. A spider had been enticed to leave the hub and run down toward a fly in the
periphery of the web while movie pictures were being taken at 18 frames per sec; the fly was
covered with quinine solution of very bitter taste. In this picture the spider just touches the
fly with a leg; in the next it has already started to withdraw. Repetition of such procedures
of movie-taking, with bitter or untreated flies, indicates that contact with one leg is suf-
ficient to transmit a chemical signal to the animal. Photograph by W. Holden.
tory of spiders rushing out to a fly in the periphery of the orb web; in several
instances the fly had been covered with quinine solution, a non-volatile sub-
stance. The pictures show clearly (fig. 5) that after barely touching the prey with
the tip of one front leg, the spider turned away if quinine was present; by
contrast it approached further and bit the prey which was untreated.
Curved, blunt-tipped hairs were found by Foelix (1970), among other
places, on the tips of the legs of web-building spiders. The tip of each hair is
open, and the lumen is innervated, so that a striking similarity exists with con-
tact chemoreceptors in insects. Such hairs would permit a spider running along a
The Web as a Means of Communication
thread to identify chemicals on the surface of the silk. While electrophysiological
proof is lacking, it is likely that the web serves also for chemical communication
between spiders.
Types of Communication in Orb Webs
Communication between Conspecifics in Early Life
In the lifetime of a spider, several striking changes can be observed in the
reaction of one animal to the presence of another animal of the same species in
one web. Maturation of the web-builder may cause the changes, alteration in
web pattern with age of the builder may be the instrument. It will be shown that
our knowledge is quite fragmentary, and that the field is wide open for experi-
mental investigations.
Several hundred young spiders emerge nearly simultaneously from an egg
cocoon. They look like small editions of adults and already at this early age,
begin to lay threads as they run. A few days after the first animals have hatched,
one can observe a sheet-like silk structure which lacks apparent geometry: it is
usually called a communal web, and is the result of all the babies’joint efforts.The
spiderlings run restlessly around on the sheet, touch each other, and lay down
additional threads. Very little is known about factors which influence the activ-
ity of the animals during this time. There exists a definite tendency to move
upward on consecutive days, and the animals appear to be attracted to each
other. If sufficient space is available, one can see, after 2—3 weeks, single spiders
in the hub of individual geometric orb webs near the communal web. Measure-
ments reveal (Witt et al, 1972) that the early orb webs look narrow-meshed,
small and very regular, and the pattern bears a striking resemblance to the adult
web.
Peters (1969) argues that the presence of a suitable frame promotes early
orb-web construction. There are some indications that lack of space postpones
construction of the first orb webs. If sufficient space is not provided in time, we
find smaller spiders falling prey to their littermates.
Spiders like Araneus diadematus radically alter their behavior toward their
siblings between the communal web stage and solitary life on the orb: they live
peacefully together on the communal web, but they attack, bite, wrap and eat
each other in the orb. The usual explanation for this change (see for example
Bristowe, 1941) goes as follows: the spiderling hatches with a yolk supply in its
abdomen. After the yolk has been exhausted, the animal feels hungry, which
releases the performed orb-web building behavior, together with the correspond-
ing behavior of trapping prey in the web. The more one thinks about such an
argument, and the closer one observes the animals during the transitional period,
the less convincing the simplicity of the causal chain becomes.
Witt
18
There is motion-picture evidence in the author’s laboratory that a spiderling
from the communal web attacks, bites, and wraps a Drosophila fly at a time
when it is still living in peace with its littermates. Even during the communal
web period it appears to be able to distinguish between a fly and other spiders. It
also looks to the observer as if in the orb-web period the violence with which a
littermate is attacked increases with time. An abrupt change in aggressiveness
does not coincide with life on the new structure. I rather believe that a change in
internal readiness, be it neural or chemical, makes a spider kill its littermate one
day, and tolerate it a day earlier. In other words, it is not the signal from one
spider to the other which determines the behavior — assuming that the signal is
different in a geometric orb and in a sheet — but rather the ‘mood’ of the
receiving animal which causes it to attack once and tolerate another time.
After the initial orb web has been built by a spider, the structural details
change in a systematic fashion from childhood to maturity. Witt et al (1972)
have shown that in Araneus diadematus the number of radii decreases, the mesh
size and irregularity of spacing increases, and the thread length and catching-area
size increase first, and later decrease statistically significantly, during the life of
an individual. There exists an age-characteristic web at all times. The obvious
question is whether the changing pattern can be simply explained by the
changing body proportions of the builder, or whether we have to look for other
variables.
Two observations speak against the first argument:
(1) if the web fine-structure were well adjusted to the body proportions of
the builder, a spider should have difficulty in using another spider’s web, if the
other individual were of different weight and size; no such difference in prey-
catching efficiency was found by LeGuelte (1969) and Richardson (1973) on
webs of spiders of different size and species.
(2) According to an extensive statistical analysis by Rawlings (Witt et al,
1972) spiders of two species Argiope aurantia and Araneus diadematus, at a time
when they showed similar body weight and leg length, built orb webs so signifi-
cantly different in several structural details, that the measures permitted classi-
fication of the builder as to its species. Body weight and leg length apparently
did not determine web fine structure.
At the present stage of our knowledge one is lead to conclude that a gradual
change in the information-receiving spider rather than a change in the trans-
mitting web structure determines alterations in behavior between conspecifics.
Communication through the Web in Mid-Life
From around the time a female builds its first orb to a few weeks after the
last molt, the spider seems to lead a lonely life in the hub of its web. It turns in
the direction in which the web is touched and shows a number of different
behaviors, obviously as a consequence of the signal it receives. As far as can be
The Web as a Means of Communication
19
established, the response to the signal also depends on the state of the animal,
and evidence shows that hunger as well as experience play a role in the regula-
tion of a spider’s reaction to web-transmitted signals. Again, the observed be-
havior is the composite result of sensory input and the inner state of the animal.
In their extensive analysis of predatory behavior in an orb-weaving spider,
Robinson and Olazarri (1971) called the first two stages prey location and prey
discrimination. Their thorough analysis of behavior units is mainly concerned
with sequences after the spider has touched the prey, and they discuss problems
of communication only marginally.
According to Robinson and Olazarri the spider is being alerted by the
impact of an object striking the web. Next it turns in the direction of the
impact, frequently plucking the radii with one or two sharp jerks of the front
legs. Plucking can continue during approach. The spider may then either bite the
prey at once, wrap it first, return to the hub, or cut and drop the prey put of the
web. Toward very large prey the spider may approach slowly, sometimes with
anterior legs raised off the web and flexed back. Little is known about the kind
of signals transmitted from prey to predator during all this time.
The alert reaction of the spider to a vibrating tuning fork touching the web
(Boys, 1880) provides evidencé that low-frequency vibrations transmitted along
radii are frequently among the first signals which reach the spider. By turning
around and plucking, the animal appears to search for further information.
Plucking may induce an immobile prey to struggle, it may serve exact; location of
the object in the web,/and it may provide preliminary information on the size
and weight of thé potential prey. The question must be asked whether a spider
can identify an object through the web (or the air) before it touches it. I believe
that it can.
The experiments of Bays (1962) support this belief: he paired one fre-
quency tuning fork with a bitter tasting fly and another frequency with a sugar-
coated animal. After a training period he could replace the flies with glass beads
of similar weights and could randomize the sequence with which he offered the
taste-frequency combination. His spiders would learn to approach, bite and wrap
one set of tasteless bemads, and cut out the other set, apparently without touching
them. If confirmed, this would show that spiders can distinguish between dif-
ferent tuning forks, and that they can avoid adverse objects in the web from a
distance. The signal transmitted by the web would provide sufficient informa-
tion to determine the subsequent type of behavior.
There is also a substantial amount of anecdotal evidence that spiders cut
dangerous bees from the web without ever touching them. They would have to
‘know’, or have learned, which signals warn of danger, and which incite approach
and attack. It is conceivable, but has never been proven, that spiders can learn
from one adverse experience about danger of certain prey. Reed et al. (1970)
found that prey-catching behavior, in contrast to web building, showed signs of
Wirt
20
plasticity: adult spiders who had never been allowed to catch prey on the web
built normal patterns, but disregarded flies in their webs.
As far as can be established from the literature and our own observations
there is no evidence that spiders in mid-life discriminate between their conspe-
cifics and other prey items in the web.
Approach of Male to Female
The mature female permits the mature male to approach through the orb
web for copulation. The elaborate ritual which precedes copulation looks to an
observer as if it has become difficult for the female web-builder to accept any-
thing but prey in its trap. There are characteristic differences from species to
species. My own observations with Araneus diadematus will be mainly men-
tioned, and some of the interesting descriptions of Grasshoff (1964) will help in
the identification of species specificity underlying communication in the web by
mating spiders.
Many weeks before maturity, males stop web building; they move around
over relatively great distance^, laying drag-lines of silk as they run. In the labora-
tory, it is nearly impossible during that time to induce them to take food, and
they stop growing. The female of the speciés, in contrast, builds large webs daily
and feeds voraciously. The female webs just before and after the last molt reach
the largest size and are built with the longest thread, if compared to webs built
at any other time in the spider’s life. At mating time the female may weigh
5—10 times as much as the male.
Frings and Frings (1968) write that the male generally finds the female by
chemical signals: ‘Her web is usually scented with a specific odor. The male
wanders about until he blunders upon the web. He is capable of determining the
species and sex, but generally not the sexual readiness of the female. If she is,
the male generally uses vibrations of the web as signals.’. The authors do not
present the evidence on which these statements are based. When the male gets to
the female web, it shows two characteristic behaviors: it fastens a ‘safety thread’
outside the female’s web which lets it swing away, and return to the web, at any
time; it plucks the thread on which the female’s front legs lie in a characteristic
rhythm. Recordings of the signals which pass through the web are to my knowl-
edge lacking, and Grasshoffs description is detailed enough to show what we
know about the communication between male and female at that time.
The male plucks a radius eight to nine times in quick succession and then
applies a strong jerk, which shakes the female visibly. After an interval of a few
seconds the process is repeated. About 5 min after the male has begun its ap-
proach, the female responds by moving toward the male, turning the abdominal
surface toward the male and shaking the two pairs of anterior legs. The male
answers with further approach, plucking with increasing speed, slowly beginning
to vibrate its whole body, interrupted by periods of immobility. Finally the male
The Web as a Means of Communication
21
jumps toward the female, holds her legs, and copulation occurs. The preliminary
approach may be repeated many times, interrupted by the swinging of the male
out of the female’s web on its security thread.
As in all other instances discussed earlier, there can be no doubt that the
web is an important substrate for the communication between the sexes. It
seems likely that chemical and vibratory signals are used, the latter being trans-
mitted through silk as well as through the air.
One interesting question addresses itself to the role of the female in the
early phases of mating; does she actively participate, sending signals back to the
courting male? I believe this to be most likely. Scarboro (personal communica-
tion) has observed in this laboratory that a web without a female is much less
attractive to a male, though some drumming and searching takes place. And even
a web with an inert weight in the hub appears to present an insufficient stimulus
for the male to continue courtship behavior. describes an Araneus
pallidus male on an Araneus diadematus web. It appears unable to arouse the
female to cooperation. Interestingly enough it took 5 h for the male to cease
approaching the wrong female; and only a few minutes later, after the male had
been removed and the proper male brought to the web, the female mated with
the male of its species.
From such a description it appears doubtful that there is a species specific
early recognition system through the web between male and female. It looks as
if, beyond a general alertness caused by the male’s drumming on the web, the
female ‘waits and sees’; the male apparently takes a long time and has to ap-
proach quite close to the female in order to be sure that it is dealing with the
proper species. The question of the function of the species-specific form of the
orb web is left wide open. Investigations have not been detailed enough, in my
opinion, to discard the hypothesis that the variation in web architecture ac-
cording to age and species has some survival value, like aiding in species isolation
or helping the male to recognize the mature female web builder. One must hope
that questions like these will be investigated in the near future.
Epilogue
There must be a difference in communication through webs between the
builders of single orbs, the subsocial spiders With groups of interconnected webs,
and the social spiders with mile-long sheet-webs ( 1972). How do
social spiders perceive the difference between members of their species and
potential prey? Krafft (1971) has identified several signals in Agelena conso-
ciata, mainly of a chemical nature, which support interindividual tolerance and
even cooperation. Little is known about the role which the web plays in commu-
nication of these spiders. To discuss this further would require too much specu-
lation for which too little knowledge exists at the present time.
Witt
22
The whole discussiorl of the web as a means of communication has suffered
from lack of exact data. Many authors have observed the phenomenon and
reported it marginally in papers which address themselves mainly to other sub-
jects. It has been difficult, sometimes impossible, to decide who should be
credited with having thought of and contributed to any of the aspects of com-
munication which were discussed here. Sometimes I have selected the earliest
known author, at another time a sentence from a recent review in order to have
one paper of each author in the reference list. This review in no way provides a
complete list of references.
Maybe the discussion of communication requires the presentation of many
different approaches, in order to lead to a final understanding of the process.
The web architecture and the chemical and physical properties of silk appeared
as important as the perceptual equipment and behavioral record of web-building
and use. At a future date it may become clear to what extent the change in web
pattern and to what extent the maturation of an animal contribute to the
extensive changes in behavior which a spider shows in reaction to signals trans-
mitted through its web.
References
Baltzer, R.: Beitràge zur Sinnesphysiologie und Psychologie der Webespinnen. Mit. Natur-
forsch. Ges. Bern 163-187 (1923).
Barth, F.G.: Bauprinzipien und adâquater Reiz bei einem Mechanoreceptor. Verh. dtsch.
zool. Ges. 66: 25-30 (1973).
Bays, S.M.: A study of the training possibilities of Araneus diadematus Cl. Experientia,
Basel 18: 423 (1962).
Blum, M.S. and Brand, J.M.: Social insect pheromones: their chemistry and function. Amer.
Zool. 12: 553-576 (1973).
Boys, C.B.: The influence of a tuning-fork on the garden spider. Nature, Lond. 23: 149-1
150 (1880).
Bristowe, W.S.: The comity of spidersM. Roy. Soc., Lond. (1941).
Finch, A.: Vibration sensitivity in an orb-weavèr. Amer. Zool. 12: 539-543 (1969).
Foèlix, R.F: Chemosensifive hairs in spiders. J. Morph. 132: 313-321 (1970).
Frings, H. and Frings, M.: in Seboek Animal communication, chap. 12, pp. 256-270 (In-
diana University Press 1968).
Gôrner, P. and Andrews, P.: Trichobothrien, ein Ferntastsinnesorgan bei Webspinnen. Z.
vergl. Physiol. 64: 301-317 (1969).
Grasshoff, M.: Die Kreuzspinne Araneus pallidus – ihr Netzbau und ihre Paarungsbiologie.
Natur Museum 94: 305-314 (1964).
Hanstrôm, B.: Fortgesetzte Untersuchungen iiber das Araneengehirn. Zool. Jb. Anat. 59:
455-478 (1934).
Jackson, R.R.: Fine structure of the thread connections in the orb web of Araneus dia-
dematus. Psyche, Heidelb. 78: 12-31 (1971).
Kaston, B.J.: The senses involved in the courtship of some vagabond spiders. Ent. Am. 16:
97-167 (1936).
The Web as a Means of Communication
23
Krafft, B.: La société d’Agelena consociata Denis, araignée sociale du Gabon. 96 e Congr.
Nat. soc. sav. Toulouse 1971, pp. 453-464.
Kullmann, EEvolution of social behavior in spiders. Amer. Zool. 12: 419-426 (1972).
Langer, R.M.: Elementary physics and spider webs. Amer. Zool. 9: 81-89 (1969).
LeGuelte, L.: Peut-on expliquer la spécificité de la toile des araignées? Rev. comp. Animal
3: 6-12 (1969).
Liesenfeld, F.J.: Über Leistung und Sitz des Erschütterungssinnes von Netzspinnen. Biol.
Zbl. 80: 465-475 (1961).
Lucas, F.: Spiders and their silks. Discovery 25: 1-7 (1964).
Lucas, F. and Shaw, J. T.B. : Comparative studies of fibroins. I. The amino acid composition
of various fibroins and its significance in relation to their crystal structure and taxon-
omy. J. molec. Biol. 2: 339-349 (1961).
Markl, H.: Leistungen des Vibrationssinnes bei wirbellosen Tieren. Fortschr. Zool. 21:
100-120 (1973).
Millot, J.: Sens chimique et sens visuel chez les araignées. Ann. Biol. anim. 22: 1—21 (1946).
Peakall, D.B.: Composition, function and glandular origin of the silk fibroins of the spider
Araneus diadematus Cl. J. exp. Zool. 156: 345-350 (1964).
Peakall, D.B.: Synthesis of silk, mechanism and location. Amer. Zool. 9: 11—19 (1968).
Peters, H.M.: Die Fanghandlung der Kreuzspinne. Experimentelle Analyse des Verhaltens. Z.
vergl. Physiol. 15: 693-748 (1931).
Peters, H.M.: Weitere’Untersuchungen Uber die Fanghandlung der Kreuzspinnen. Z. vergl.
Physiol. 19: 47-67 (1933).
Peters, H.M.: Maturing and coordination of web-building activity. Amer. Zool. 9: 223—227
(1969).
Reed, C.F.; Witt, P.N.; Scarboro, M.B., and Peakall, D.B.: Experience and the orb web.
Develop. Psychobiol. 3: 251-265 (1970).
Richardson, C.E.W.: Specificity of the spider-web pattern and its efficiency in trapping
prey. Fed. Proc. 13: 4 (1973).
Robinson, M.H.: Predatory behavior of Argiope argentata Fabricius. Amer. Zool. 9:
161-173 (1969).
Robinson, M.H. and Olazarri, J.: Units of behavior and complex sequences in the predatory be-
havior of Argiope argentata Fabricius. Smithsonian Contr. Zool. Wash. 65: L-36 (1971).
Salzmann, L. and Witt, P.N. : Life on a thread. An introduction into the study of behavior
(Blue Flower Films, Raleigh, N.C. 1973).
Seyfarth, E.-A. and Barth, F.G.: Compound slit-sence organs on the spider leg: mechanore-
ceptors involved in kinesthetic orientation. J. comp. Physiol. 78: 176—191 (1972).
Walcott, C.: A spider’s vibration receptor: Its anatomy and physiology. Amer. Zool. 9:
133-144 (1969).
Warwicker, J.O.: Comparative studies of fibroins. II. The crystal structure of various fi-
broins. J. molec. Biol. 2: 350-362 (1960).
Warwicker, J.O.: Comparative studies of fibroins. V. X-ray examination of chemical resis-
tant fractions of some silk fibroins. Biochim. biophys. Acta 52: 319—328 (1961).
Webster’s third new international dictionary (Miriam Co., Springfield, Mass. 1965).
Witt, P.N.: Behavioral consequences of laser lesions in the central nervous system of Araneus
diadematus Cl. Amer. Zool. 9: 121—131 (1969).
Witt, P.N.; Reed, C.F., and Rawlings, J.O.: Ontogeny of web-building behavior in two
orb-weaving spiders. Amer. Zool. 12: 445-454 (1972).
Dr. P.N. Witt, Department of Human Resources, Division of Mental Health Services, Re-
search Section, Raleigh, NC 27611 (USA)