Images Collection
View this article in Search Friendly Plain Text
NOTE: This plain text article interpretation has been digitally created by OCR software to estimate the article text, to help both users and search engines find relevant article content. To read the actual article text, view or download the PDF above.
Jackson, R. R. 1974. Effects of d-amphetamine sulfate and diazepam on thread connection fine
structure in a spider’s web. J. Arachnol. 2:37-41.
EFFECTS OF D-AMPHETAMINE SULFATE AND
DIAZEPAM ON THREAD CONNECTION FINE
STRUCTURE IN A SPIDER S WEB1
Robert R. Jackson2
Division of Research
North Carolina Department of Mental Health
Raleigh, North Carolina 27611
ABSTRACT
Dextro-amphetamine and diazepam were administered orally to the spider Araneus diadematus.
Thread connections from the spider’s orb web were examined by photomicroscopy. Amphetamine
altered fine structure but diazepam did not. Results are discussed in terms of disturbed spinning
behavior.
INTRODUCTION
When a spider constructs its web, it produces a record of its web-building behavior
which can be readily measured and quantified. Through the spider’s web, we have a very
sensitive measure of drug effects (Witt and Reed, 1968), and motor coordination distur-
bance has been indicated for a number of drugs. Previous studies, using such measures as
radial angle regularity, spiral regularity, and total thread length, have dealt with the gross
structure of the web. However, the spider does not merely place the threads in space
according to a certain configuration, but fastens threads to each other at over 1000 distinct
locations. The complex fine structure of thread connections has been described (Jackson,
1971), but the details of how the spider forms a thread connection are not known.
Probably, fine coordination of body, leg, and spinneret movements plus coordinated
functioning of several glands are involved. The present study investigated the possibility
of using thread connection fine structure as a measure of drug effects on fine motor
coordination.
Dextro-amphetamine, a central nervous system stimulant, was chosen because studies
on web gross structure indicate that it causes motor coordination disturbance (Witt,
Brettschneider, and Boris, 1961), and therefore it would be a likely candidate to produce
alteration in thread connection fine structure, if the fine structure is sensitive to drugs.
Diazepam, a tranquilizer, causes curtailment of thread expenditure with no effect on silk
synthesis, but at the concentration used in the present study, motor coordination distur-
bance is not indicated (Reed and Witt, 1968). This drug was chosen because of the
possibility that fine motor coordination disturbance might occur when a drug is
administered to a spider and yet not be detected in the gross structure of the web.
‘Supported in part by a grant from Hoffmann-LaRoche, Inc., Nutley, New Jersey, and by Grant
number GB-6246 from the National Science Foundation to P. N. Witt.
2 Present address: Department of Zoology, University of California, Berkeley, California 94720.
37
THE JOURNAL OF ARACHNOLOGY
METHODS
Adult females of Araneus diadematus Clerck were administered drugs in the previously
determined effective doses of 100 mg/kg for diazepam (Reed and Witt, 1968) and 300
mg/kg for d-amphetamine sulfate (Witt et al., 1961). The pure substance was dissolved (or
suspended) in sugar water and given to the spider in a volume of 100 jjl! with a Hamilton
microsyringe. The spider drank the drop from the syringe as she sat on her web. Drugs
were administered approximately 24 hours (diazepam) and 12 hours (amphetamine)
previous to the spider’s next web-building period. Once it was determined that the spider
had consumed the drug, the old web was destroyed. Sticky spiral to radius connections
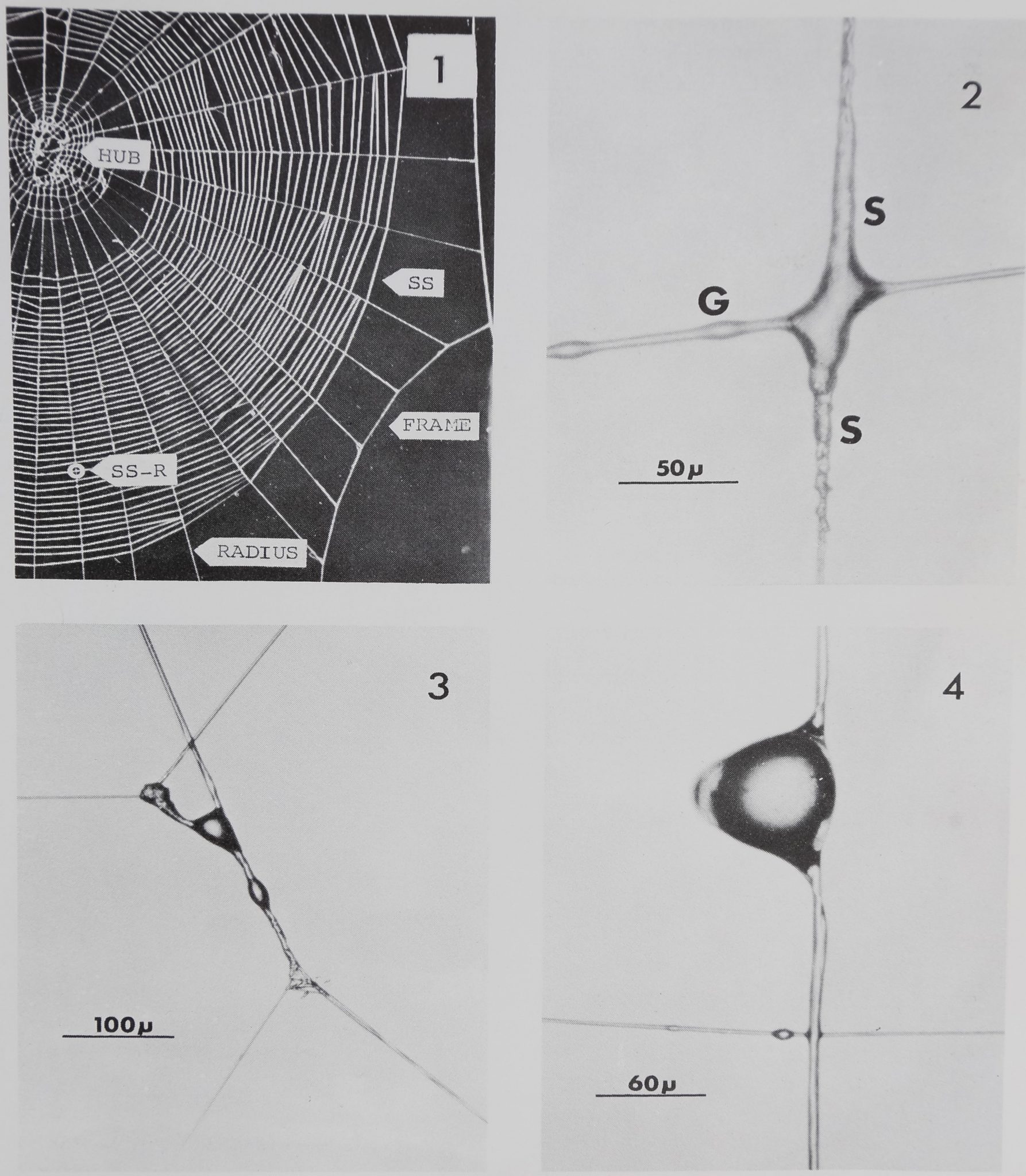
(SS-R’s) from the spider’s next web were examined. This is the most abundant type of
connection in the spider’s web (Fig. 1). Thirteen SS-R’s for amphetamine (amphetamine
SS-R’s) from 2 webs built by different spiders and 6 SS-R’s for diazepam (diazepam
SS-R’s) from 2 webs built by different spiders were compared with 49 normal SS-R’s
from 15 webs of 8 spiders which were not given drugs. These normal SS-R’s were
described in a previous study (Jackson 1971). Procedures for maintaining the spiders and
obtaining photomicrographs of thread connections were identical to those previously
described (Jackson, 1971 ; Witt, 1971).
RESULTS
The fine structure of a normal SS-R is shown in Fig. 2. The appearance of a sleeve, i.e.,
a rougher appearing area on a radius or spiral thread near the junction, was one of the
most characteristic features of a normal SS-R, but these were less abundant at
amphetamine SS-R’s. At normal and diazepam SS-R’s there was always a sleeve on the
radius at the side of the connection closer to the frame of the web, but 4 out of 13
amphetamine SS-R’s did not have a sleeve at this location (Fig. 3 and 4). This was
significantly different from normal SS-R’s (X2 = 11.422, df = 1, P< 0.05). There was a
sleeve on the radius at the side of the connection closer to the hub of the web for 23 out
of 49 normal SS-R’s and 2 out of 6 diazepam SS-R’s, but there was a sleeve at this
location for only 1 out of 13 amphetamine SS-R’s. Diazepam and normal SS-R’s were not
significantly different at the 0.05 level (X2 = 0.039, df = 1, n. s.). Amphetamine SS-R’s
were significantly different from normal SS-R’s(X2B= 5.118, df=l,P<0.05). The thick?
ness of spiral threads varied over a greater range at amphetamine SS-R’s. The maximum
thickness at normal and diazepam SS-R’s was 5 ju; for amphetamine SS-R’s it was 7 /!. The
minimum for normal and diazepam SS-R’s was 3 ju; for amphetamine SS-R’s it was 2 ju.
Three amphetamine SS-R’s had globules on the radius (Fig. 4), which appeared to consist
of the same substance as the globules on the spiral. Globules were not found on radii at
normal and diazepam SS—R’s. Due to the differences described here plus others which are
less easily described, the overall appearance of amphetamine SS-R’s was often rather
bizarre when compared to normal and diazepam SS-R’s (Fig. 3 and 4). For statistical
analysis, no distinction was made between data from different webs for a given treat-
ment. However, the effects reported for amphetamine occurred in both webs, and there
were no obvious differences between the two webs of spiders treated with diazepam or
among the webs of normal spiders.
DISCUSSION
This study has shown that, like the gross structure of the web, the fine structure of
thread connections can be used as a measure of drug effects. Consistent with studies on
Fig. 1.—Lower right quarter of a normal web of Araneus diadematus. Note that sticky spiral to
radius connections (SS-R) are the most common type of connection in the web. SS: sticky spiral.
Fig. 2.—Sticky spiral to radius connection from normal web. In photomicrograph, spiral runs
horizontally, radius runs vertically, and upward is toward frame of web. Note sleeves (S) on radius and
globules (G) on spiral.
Fig. 3.—Bizarre appearing sticky spiral to radius connection produced by amphetamine treated
spider.
Fig. 4,—Sticky spiral to radius connection produced by amphetamine treated spider. In photo-
micrograph, spiral runs horizontally, radius runs vertically, and upward is toward frame of web. On
radius, note large globule and absence of sleeves.
THE JOURNAL OF ARACHNOLOGY
the gross structure of the web, there is evidence of motor coordination disturbance for
amphetamine in the fine structure of thread connections, but at low doses not for
diazepam. However, in the case of diazepam, the small sample size should be considered.
We would like to know precisely what components of the spider’s spinning behavior
were deleted or altered in those spiders which were administered amphetamine. This
would be useful not only for understanding how drugs affect behavior but also in under-
standing how the normal spider fastens threads in the web. The normal spider goes
through a fixed sequence of leg and body movements as it constructs a SS-R (Jacobi-
Kleemann, 1953). Frame by frame motion picture analysis has shown that this sequence
is still there after amphetamine has been administered (Peters, 1950), but the rhythm is
altered, the spider sometimes moving significantly more slowly or more rapidly than usual
(Witt, Reed, and Jackson, 1972). Altered rhythm may be at least partly responsible for
the altered thread connection fine structure. For example, the plasticity of substances
from the silk glands, either the silk itself of special glue substances (Jackson, 1971), may
decrease radidly after leaving the spinnerets. Stretching movements slightly earlier or later
that usual may produce very different effects. However, there is another possibility that
needs to be considered. The spinnerets are capable of complex movements (Wilson,
1969), and the spider has 6 different types of silk glands (Peakall, 1969); but the fine
motor coordination of spinneret movement and gland functioning during thread connec-
tion construction has not been analyzed in detail. Perhaps amphetamine produces distur-
bances at this level.
Some amphetamine SS-R’s resembled artificial thread connections (Jackson, 1971) in
general appearance, in lacking sleeves, and, in some cases, by having a globule on the
radius. The artificial thread connections were made by taking a sticky spiral thread from
the web and simply placing it across a radial or frame thread and, in some cases, stretch-
ing the spiral thread with an oil coated insect pin (Jackson, 1971).||hese connections
were never securely fastened. Points at whiA threads touch but are not securely fastened
are rare in the vertical web of A. diadematus; however such points are frequent in the
horizontal orb web of Uloborus diversus (Eberhard, 1972). Perhaps amphetamine treated
spiders sometimes do something similar to making an artificial connection, i.e. placing the
threads and jerking the spiral but somehow failing to properly perform the more subtle
behavior involved in fastening the threads.
ACKNOWLEDGEMENTS
The assistance of Dr. Peter Witt, Dr. Roy Caldwell and Mrs, Mabel Scarboro is grate-
fully acknowledged.
LITERATURE CITED
Eberhard, W. G. 1972 The web of Uloborus diversus (Araneae: Uloboridae). J. Zool., London
166:417-465.
Jackson, R. R. 1971. Fine structure of the thread connections in the orb web of Araneus diadematus.
Psyche 78:12-31.
Jacobi-Kleemann, M. 1953. Ueber die Lokomotion der Kreuzspinne Aranea daidema beim Netzbau
(nach Filmanalysen). Z. vergl. Physiol. 34:606-654.
Peakall, D. B. 1969. Synthesis of silk, mechanism and location. Amer. Zool. 9:71-80.
Peters, H. M. 1950. Institut fuer Film und Bild in Wissenschaft und Unterricht (Goettingen,
Germany, 1950), Hochschulfilm C 596.
EFFECTS OF D-AMPHETAMINE SULFATE AND DIAZEPAM ON SPIDER WEBS
41
Reed, C. F. and P. N. Witt. 1968. Progressive disturbance of spider web geometry caused by two
sedative drugs. Physiol. Behav. 3:119-124.
Wilson, R. S. 1969. Control of drag-line spinning in certain spiders. Amer. Zool. 9:103-111.
Witt, P. N. 1971. Instructions for working with web-building spiders in the laboratory. Bioscience
21:23-25.
Witt, P. N., L. Brett schneider, and A. P. Boris. 1961. Sensitivity to d-amphetamine in spiders after
iproniazid and imipramine. J. Pharmacol. Exptl. Therap. 132:183-192.
Witt, P. N. and C. MReed. 1968. Altered web patterns, p. 39-73. In P. N. Witt, C. F. Reed, and D. B.
Peakall, A spider’s web. Springer Verlag, New York.
Witt, P. N., C. F. Reed, and R. R. Jackson. 1972. Rhythmic activity of the web-building spider, and
its disturbance by dextro-amphetamine. Fifth Intern. Congr. Pharmacol., July 23-28, 1972,
Abstr. Volunteer Papers, Abstr. 1525:255.