Images Collection




View this article in Search Friendly Plain Text
NOTE: This plain text article interpretation has been digitally created by OCR software to estimate the article text, to help both users and search engines find relevant article content. To read the actual article text, view or download the PDF above.
Am. Zoologist, 12:445454 (1972).
Ontogeny of Web-building Behavior in Two Orb-weaving Spiders
Peter N. Witt
Research Division, North Carolina Department of Mental Health,
Raleigh, North Carolina 27611
John O. Rawlings
Department of Statistics, North Carolina State University, Raleigh,
North Carolina 27607
AND
Charles F. Reed
Department of Psychology, Temple University, Philadelphia, Pennsylvania
19122
synopsis. Spiders of similar mass but of two different species, Araneus diadematus Cl.
and Argiope aurantia L., build webs which hâve similar geometric patterns, but which
differ in relative proportions and thread number. Within species, webs may also differ
reliably, presumably on a genetic basis. Fine detail of the web undergoes change
throughout the lifetime of the spider, different for male and female; these changes are
not simply responses to the growing weight of the spider. Relatively simple restraints,
such as availability of material for thread, result in adaptations of web-building which
may at first appear to be based upon building experience, but which are in fact
independent of it.
It is possible to conceive of the form of
the orb web as the necessary outcome of
the physical restrictions imposed upon a
thread-placing apparatus (the spider).
The threads must support a spider of a
certain mass, endure violent movement,
and remain a functional snare for many
hours. It would be expected that webs of
the same class, e.g., vertical cartwheel orbs,
would resemble each other in form.
At the same time, it would be expected
that there would be variations in the web
The support of part of this work through grant
number GB-6246 from the * National Science
Foundation, statistical work by J. Chappell, and
laboratory assistance by M. Scarboro are gratefully
acknowledged by the authors.
The contribution of the second author was sup-
ported in part by Public Health Service Grant GM
11546 from the Division of General Medical
Sciences.
Joint contribution of the Division of Research,
North Carolina Department of Mental Health and
the Department of Statistics, North Carolina State
University, Raleigh, North Carolina. Listed as Pa-
per No. 3777 of the Journal Series of the North
Carolina State University Agricultural Experiment
Station, Raleigh, North Carolina.
set by physical dimensions of the spider,
e.g., weight. Beyond the physical demands
for placing a thread network, there would
be functions particular to the developmen-
tal stage, sex, or species of the spider — all
somehow innately programmed into execu-
tion of web-building.
The cartwheel webs of different species
may resemble each other because of the
engineering requirements of structure and
thread, or because of similarity of central
nervous system programming. There are
particular features such as the stabilimen-
tum of Argiope and the open sector of
Zygiella which obviously distinguish the
webs, but the fundamental geometric
forms are very similar. To the degree that
these common features occur, the form of
the orb web may be viewed as a convergent
adaptation of behavior to the problem of
arraying threads in space for trapping air-
borne prey (Figs. 1, 2).
Nevertheless, the features of the web
change throughout the life of the spider. It
is useful to examine these changes for
clues to the relative contributions of innate
445
446
Witt, Rawlings, and Reed
FIG. 1. A web of an adult female Argiope aurantia istic differences in shape (width over length), fine
on the left and an adult female Aran eus diade- structure and relative size of center area in the
matus, as they are rebuilt nearly every day in the webs of the two species (compare to Table 3).
50 x 50 cm laboratory frames. Note the character-
F1G. 2. The webs of an adult female Zygiella-x- structure, but differ through the free sector with
notata (on the left) and a subadult female Neos- signal thread, which only Zygiella builds. Compare
cona hentzii are very similar in shape and fine these webs to the two in Figure I.
Ontogeny of Web-building Behavior
447
FIG. 3. Webs built by female Araneus diadematus
in the laboratory frames at ages 27, 120, and 437
days; all three photographs are enlarged to the
same scale, the vertical white lines being originally
spaced 20 mm apart. Note the characteristic
features like small meshes, relatively short thread.
and small catching area in the web built by the
young spider, y|s compared to the larger meshes,
longer thread, and large catching area in the web
of the adult. The old female builds a rather coarse
meshed and irregular web. (Compare to Figs.
4, 5.)
movement programs, physical restraints,
and perhaps even the experience of the
spider in building as an example of the
ontogeny of a specific behavior.
The webs of young and old animals of
two different species of spider were mea-
sured daily from hatching to death. All
observations were performed in a laborato-
ry kept in a cycle of long warm days and
short cool nights throughout the 8- to
16-month lifespan of the animals. Individ-
ual spiders were kept in aluminum cages
large enough to impose no limitation on
the web proper, i.e., on the catching zone
as distinct from the framing and mooring
threads (Witt, 1971). For very young ani-
mals, a small wooden frame was also
placed in the cage; the animals were not
restricted to the frame.
Webs were photographed shortly after
they were built each day and were then
collapsed by the experimenters; as a rule,
the silk was ingested by the spiders and
new webs were built the following day.
The collapsing procedure does not affect
frequency of building; even when undis-
turbed, the spider dismantles and ingests
the web (Breed et al., 1964). We assume
that the course of events for laboratory ani-
mals is essentially similar to that for ani-
mals living outdoors.
The web of Araneus diadematus is typi-
cal for vertical orb webs: a structure with
many radial threads on which is wound a
spiral, plainly continuous only toward the
center of the web, a more or less isolated
hub or platform at the center, and a very
variable scaffolding which supports the
web proper. While retaining this general
structure, the web varies in detail from day
to day. Two webs of the same overall size
may differ in the areas devoted to the cen-
tral or spiral zones. Threads may be very
evenly spaced or relatively irregular with
large open areas. The number of spiral
and radial strands, the relatively oval
shape of the catching zone, and the total
length of thread show different values over
time and between spiders. Statistical group-
ing reduces the variations of short time-
course and provides an impression of long-
term development of web characteristics
(Witt and Reed, 1965).
Age. Some of the age-related changes in
Witt, Rawlings, and Reed
ad^o
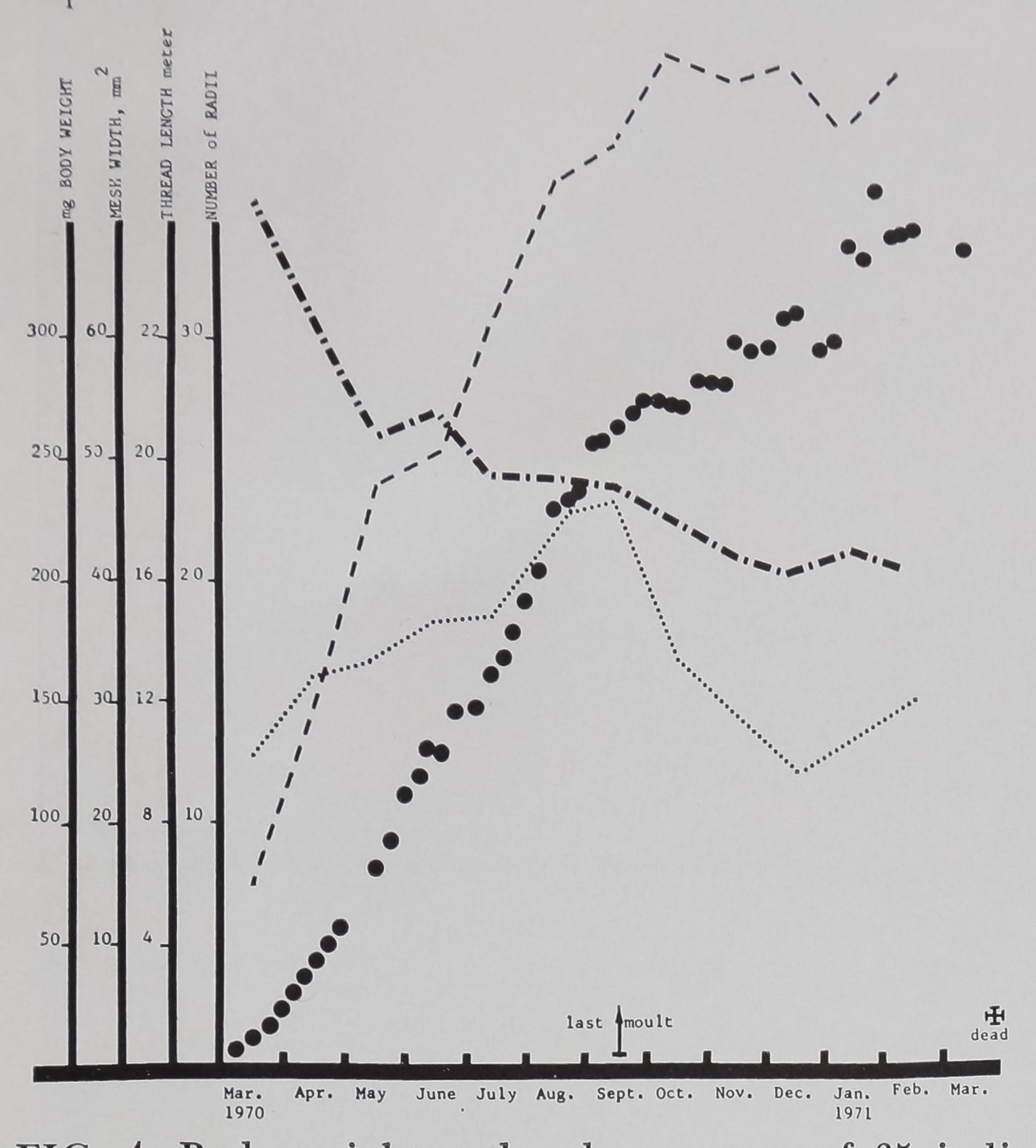
FIG. 4. Body weight and web measures of 35 indi-
vidual female littermates in Family 1 of Aran eus
diadematus Cl., hatched in the laboratory on the
25th of February 1970. Black, full circles represent
mean body weight once a week; dashed line con-
nects monthly means of mesh width in all webs
measured in that month; dotted line connects
means of all thread lengths; dash-dotted line num-
ber of radii. Note the various combinations of web
characteristics at different times of life, and their
relationship to the time of the last moult or of
reaching sexual maturity. (Compare to Fig. 3.)
webs of a single A. diadematus female are
apparent in Figure 3. A more complete
record of web-building ontogeny was ob-
tained from a single cocoon with eggs
which hatched in the laboratory in late
February (Fig. 4).
The hatchlings were presumably the
offspring of a single mating. Over 100 ani-
mals emerged from the cocoon, and over
the next two weeks traveled short distances
back and forth from the cocoon, laying
threads and creating a structure with no
apparent ordering, resembling a silken
sheet. In early March, individual animals
drew apart and constructed geometric orb
webs which showed all of the characteristic
features of the adult web: hub, radii,
catching spiral, and frame. At this point,
the animals were transferred to individual
cages, where they received food and water
twice a week. Webs were photographed
once a week, after they had been cut the
day before, and the spiders were weighed
weekly. Similar measures were taken for
cocoons hatching shortly before and after
this cocoon in order to confirm the gener-
ality of the findings for the species.
The mean weight of female A. diadema-
tus increases with age in progressive fash-
ion. Since the graph for mesh size (Fig. 4)
follows the same general course, weight
would seem to be one of the determinants
of the texture of the web. The measure
“mesh width” refers to the fineness of the
network formed in the catching zone by
the radial and spiral threads (Witt et al.,
1968). A relatively large web with few radi-
al or spiral turns would show a coarse
texture. Given equal web size, the greater
the number of radial and spiral turns,
the finer the texture — the threads being
placed at uniform distances. An index
of the regularity of that spacing is pro-
vided by computing the variance of the
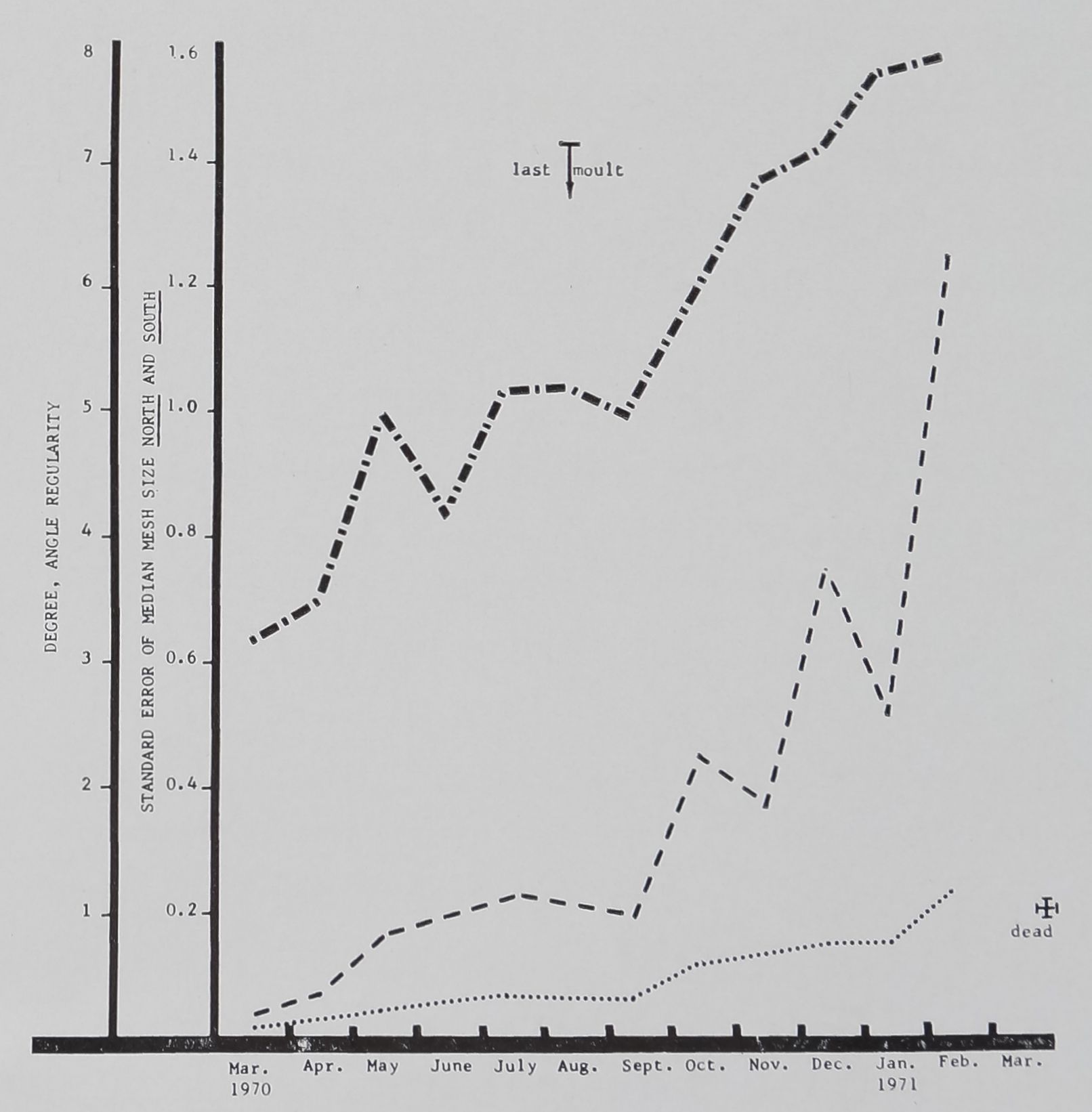
FIG. 5. Regularity measures of all webs built in
each month by 35 female littermates hatched in
the laboratory on the 25th of February 1970.
Dotted line connects means of standard error of
median mesh size South; dashed line the means of
the same measure in the North part of the web;
dash-dotted line the means for angle regularity.
Note that all measures indicate increase in irregu-
larity with increase in age, while webs get increas-
ingly wide-meshed. (Compare to Figs. 3, 7.)
Ontogeny of Web-building Behavior
449
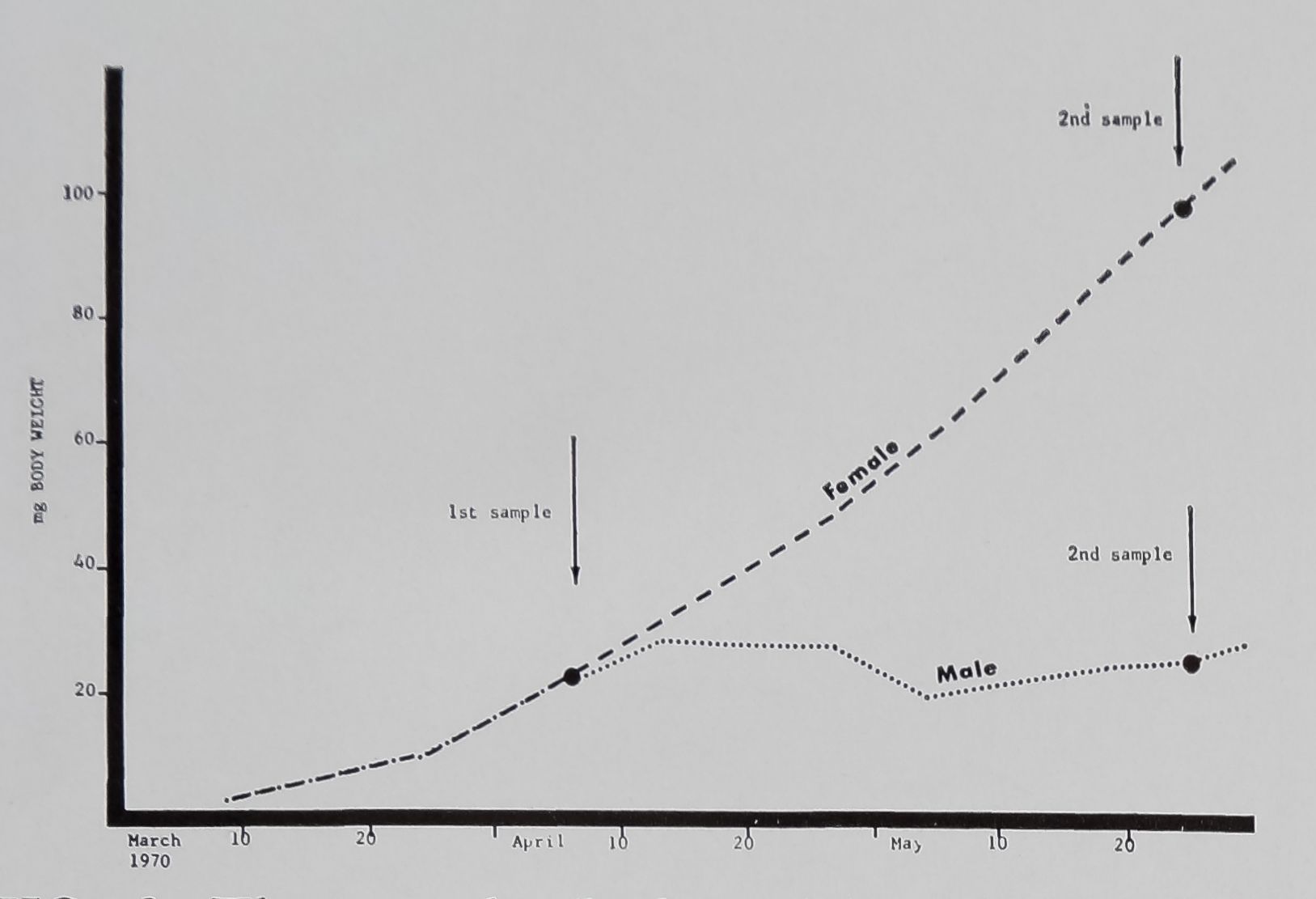
Aft.*»
FIG. 6. The growth (body weight) of 35 female
Araneus diadematus is compared to that of 30
males (littermates) during the first 3.5 months of
their lives; vertical arrows indicate days on which
webs were measured {see Table 1) . Note the
cessation of growth in males by the middle of
April as compared to the continuouBgrowth of
females; by the end of May, females weighed about
5 times as much as males.
mesh sizes in several portions of the web
(SE median mesh size). In the case of this
sample of spiders, the web becomes progres-
sively coarser throughout life until some
time after the final molt, when there is also
great variability in weight because of the
production of eggs. In contrast to Eber-
hard (1971), who reported differences be-
tween “senile virgin’s webs and senile non-
virgins” for Ulob or us diver sus, we found in
21 old Argiope aurantia female litter-
mates no correlation between changes in
web measures and deposition of the egg-
cocoon.
Wiehle (1927) states that he always
found more radii in webs of young
Araneus diadematus than in those of old
animals. This agrees with the steady de-
crease in number of radial threads shown
in Figure 4, and is in keeping with the
progressive openness of the mesh. Figure 5
shows three direct measures of regularity
of thread placement: the central angles
formed by the radial threads become pro-
gressively more variable in size, while the
standard error of the mesh also increases,
especially in the lower portion of the web.
Thread length — the total linear extent
of silk used in building the web -Reaches
a peak value at about the time of the last
molt and then declines. Previous experi-
ments (Christiansen et al., 1962) have
shown that the threads of the web become
thicker as the weight of the spider is in-
cMBed (by attaching a weight to the ab-
domen). The same amount of silk may be
employed, but the length of thread is cur-
tailed.
The general effect — which the webs of
Figure 3 typify —is that young A. diade-
matus females build small,. fine-meshed
webs, mature animals large wide-meshed
webs, and old animals coarse and some-
what irregular large webKÿ
Despite this general picture, it can be
shown that the regularity of the web is not
necessarily a Bonsequence of the size of
the spider. Males do not grow to the same
size as females (Fig. 6), and although fre-
quency of building drops drastically for
males at an early age, they do build a
table 1. Mean weight and web measures of male and -f^aZljAlpieus diadematus littermates
iy2 and 3 months after hatching.
Early^webs Lafe webs
Measurements v^3e4 males 8 males 20 females
mg Body weight 23.69 26.69 26.41 116.30*
mm Leg length cm2 Spiral area 7.5f , 7.75 9.75 11.95
19.47 19.18 14.98 33.40*
cm2 Center area 1.64 1.77 ‘ 1.58 3.15*
m Thread length 11.21 10.98 8.94 14.64<*>
mm2 Mesh mean 23.58 26.30 25.44 63.14*
SE med. mesh size South 0.021 0.022 0.036* 0.033*
Note that late female webs differ significantly from other webs in 4 measures; while late male
and female webs differ from all early webs in the regularity measure.
* Significant difference below P 0.01 ; analysis of variance.
<#) Significant difference below P 0.05; analysis of variance.
450
Witt, Rawlings, and Reed
FIG. 7. The upper two webs were built by two
female Araneus diadematus, the lower by litter-
mate males. When building the two webs on the
left, the spiders were 1.5 months old, and the webs
on the right were built when they were 3 months
old. (Compare to Fig. 6) . All webs are reproduced
sufficient number to permit comparison
with the late webs of females.
Table 1 contains the body and web mea-
sures for A. diadematus males and females
at^ ithe same scale. The webs show characteristic
features which identify the builders; relatively high
irregularity in thread spacing on the right by
the more mature animals, and a comparatively
large web built with a long thread by the large
female. Compare to mean measures in Table 1.
at two periods of life: in April when mass
and leg length are equal for both, and at
the end of May, when sexual maturity has
been reached and the mean body weight of
Ontogeny of Web-building Behavior
451
the female spiders has increased fourfold.
The males remain at approximately the
same mean weight.
Four measures of the web (spiral area,
central area, median mesh size, and SE of
median mesh — the last a measure of vari-
ance) show significant differences between
the webs of mature females and all other
entries, with the important exception of
the variance of the mesh size. In that case,
the late male and female spiders show
equal irregularity of spacing. A measure
not shown in the table S regularity of
spacing of the spiral thread — also de-
creases significantly for the older animals
(see Fig. 7). These findings suggest that
regularity of spacing is related to matura-
tion rather than simply to growth in size.
(As examination of the table shows, it is
not possible to relate the regularity mea-
sures simply to leg length: the older male
values occupy a position between the leg
lengths of the young spiders and the leg
length of the older females.)
It does not appear that this relative de-
cline in web regularity is of a magnitude
which decreases the efficiency of the web
for catching prey. The spider must move
rapidly in any case to seize insects which
are entangled by the threads; the impor-
tant factor for trapping seems to be the
area covered by the web, within certain
limitation of finenesljof the mesh.
Experience. It is often conjectured that
experience in web-building serves to modi-
fy the form of the web throughout the life
of the spider. We have examined this hy-
pothesis by differential treatment of spi-
derlings from the time of hatching to
several months of age (Reed et al., 1970).
Two groups of spiders were kept in nar-
row glass tubes for 7 or 28 weeks while
littermate controls were reared in labora-
tory cages in which they could build full-
sized webs daily. The spiders in the tubes
were unable to build though they did
emit thread as they moved about in the
cramped space. When the confined spiders
were released, they built webs significantly
smaller than those of their unconfined lit-
termates. In several days, they were con-
structing webs of normal size.
However, adult spiders with ample
building experience also showed reduction
in size of the web if confined for only
three weeks, suggesting the presence of a
limiting factor other than experience. If
the silk glands of the confined animals
were emptied by pulling silk from the
spinnerets each of three days prior to re-
lease, the webs built after release were of
normal size and detail, if compared to lit-
termates of the same age. Finally, measure-
ment of the rate of silk production in the
glands showed that failure to empty the
glands had a slowing effect upon synthesis,
and led us to the conclusion that relative
scarcity of silk rather than of experience
accounted for the small webs of the
confined spiders. Inasmuch as the webs did
not differ in other respects, it appears that
experience plays no role in construction
of the age-specific web. This conclusion of
course does not necessarily apply to other
aspects of the spiders’ behavior.
Genetic differences. Long familiarity
with orb web geometry within a single spe-
cies encourages the impression that there
are individual differences in the form of
webs and webs characteristic of littermates
of single cocoons. There are signs of indi-
vidual preference for hub position for in-
stance (Witt, 1956, p. 20) and family char-
acteristics in size (Reed et al., 1970).
A recent study (Rawlings and Witt,
1972), using webs spun by A. diadematus
females of nearly equal weight, showed a
high repeatability for many characteristics
of webs spun one week apart by the same
spider. This, of course, reflects the joint
effects of the unique genetic control of web
building behavior for each spider and the
composite of all environmental factors
influencing the development of their indi-
vidual behavior patterns. Most notable for
the magnitude of their repeatabilities (>
0.5) were the size measures: radii number,
number of spirals in the four cardinal di-
rections, frame area, spiral area, center
area, and thread length.
Full-sib correlations from the same study
provide measures of the similarity of the
452
Witt, Rawlings,, and Reed
table 2, Mean body weight of females of two species of spiders at the time at which they built
webs which were analyzed for similarity in web measures.
No. individuals Weight in mg at time of building of
Species Family in family First web Second web
Araneus diadematus I 29 127.1 139.5
II 18 121.7 137.0
III 24 119.1 136.5
IY 4 120.5 138.8
Y 4 115.8 125.3
VI 1 116.0 135.0
YII 11 121.1 136.7
VIII &1|S| 130.0 150.0
Argiope aurantia I 13 104.2 126.3
II 18 126.3 132.7
behavior patterns of the full sibs which
should be influenced primarily by the gen-
etic factors and to some lesser degree than
with repeatability by the common cocoon
environment of the full sibs. If one could
assume no common environmental influ-
ence of the cocoon environment on the
web building behavior of the family mem-
bers, the full-sib correlation measures diff-
ering from zero would reflect the presence
of genetic variation for that aspect of be-
havior and imply some degree of genetic
control. Again, traits primarily measuring
size of web showed the highest full-sib
correlations. The fineness, or intricacy, of
the web as measured by medium mesh size
and mesh width were next in importance.
Only five of the 27 traits failed to show
significant full-sib correlations.
To extend this analysis one step further,
two families of Argiope aurantia were
sampled in a manner similar to that
above. Measurements were taken on webs
spun two weeks apart by females of nearly
equal weight. The information from Ar-
giope was combined with the A. diadema-
tus data for further analysis. All measure-
ments of basic features common to the
webs of both species were used; stabilimen-
ta of Argiope were not included (Table
2).
Two types of analyses were used. The
univariate analysis of variance was used to
determine the importance of species differ-
ences in comparison with differences be-
tween families within species. Any indica-
tion of species differences must be attribu-
table to a greater genetic similarity of
families within species than of families in
different species since there should be no
common environmental components con-
tributing to the similarities of different
families. The second analysis was a multi-
ple discriminant analysis to determine the
efficacy with which individual spiders
could be classified into the correct family
and the correct species solely on the basis
of their web building behavior. Again, this
is used only as an indicator of the degree
of the innate differences in web-building
behavior, presumably reflecting differences
table 3. Web measures showing significant (p <
O.of differences betwWm means for two species of
orb-weavers (eight families of Araneus diadematus
and two of Argio® aurantia, all females of nearly
equal body weight) ; means are adjusted for re-
ma^fhsihîg body weight differences.
Araneus Argiope
Web measures diadematus aurantia
Radius ratio 0.67 0.95
(north over south)
Radius ratio 0.73 0.89
(east over west) Frame area (mm2) 12905 17051
Spiral area (mm2) 46459 68683
Center area (mm2) 3550 7646
Center over spiral 0.085 0.130
area (ratio) Mesh size width (mm2) 58 88
Relative deviation of 0.38 0.45
spirals (south)
Median mesh size— 51 112
north (mm2)
Standard error of median 0.15 0.30
mesh size, north
Ontogeny of Web-building Behavior
453
table 4. Summary of multiple discriminant clas-
sification of webs of Araneus diadematus and Ar-
giope aurantia. Discriminant based on first five
families listed, individuals of remaining five fam-
ilies (all lenown to be A. diadematus) classified ac-
cording to the discriminant function.
Family No. of spiders classified into family
A. diadematus A. aurantia
1 2 3 4 6
1 24 2 3 0 0
2 2 16 0 0 0
3 2 1 21 0 0
4 0 0 0 12 0
5 0 0 0 1 17
6 4 0 0 0 0
7 3 0 1 0 0
8 1 0 0 0 0
9 6 0 5 0 0
10 0 0 1 0 0
in genetic programming.
Based on the univariate analysis, ten
measures significantly (p < 0.05) discrimi-
nated between the two species (Table 3).
In all cases, species differences were ad-
justed for spider weight differences before
being tested.
Considering only the shared features of
the web, the data show that, despite simi-|
lar bodily characteristics, the webs of the
two species differ in important ways (see
Fig. 1). A. aurantia appears to build larger
webs than does A. diadematus, the center
area, comprising hub and a free zone,
being twice as large. The hub is placed
more nearly in the geometric center of the
web and the texture is larger meshed and
more variable, particularly in the north-
ern half, in aurantia than in diadematus.
The multiple discriminant analysis
(using families as categories) was run in
two ways. Data from all ten families were
used in one analysis to construct the
discriminant function. In this case, even
though the analysis was terminated when
only six measures had been incorporated
into the function, 70% of the individual
spiders were correctly classified by the
discriminant as to family and 99% were
correctly classified as to species, i.e., one
individual in Family II of A. aurantia was
misclassified into Family II of A. diadema-
tus. The second analysis utilized data from
only five families to construct the discrim-
inant (the three families of A. diadema-
tus having the largest number of spiders
and the two families of A. aurantia), and
then the twenty-one spiders of the other
five A. diadematus families were classified
according to the discriminant. In this case,
90% of the spiders used in computing the
discriminant were correctly classified as to
family and all were correctly classified as to
species. Further, all 21 of the other spi-
ders were correctly classified as being mem-
bers of A. diadematus (Table 4).
The relatively low rate of error in clas-
sification by family and by species encour-
ages the speculation that there are family
and species differences, and since experi-
ence seems to be unrelated to the construc-
tion of the web, that these differences have
a genetic basis.
The pattern of the orb web, which can
be regarded as a record of a significant
portion of the builder’s observable behav-
ior, changes in a characteristic way during
the life of the spider. Such changes are
independent of the spider’s prior building
experience. There are some features of the
web, such as mesh size which seem to be
related to growth in size. There are others,
such as the regularity of those meshes,
which seem to be functions of maturation:
the late female webs are measurably differ-
ent in size only from the late male webs.
Other features of the web, thread length
for instance, are sensitive to material
requirements such as the availability of
silk.
While the general pattern of the orb
web may be viewed as a successful adapta-
tion to catching airborne prey Sand per-
haps may be an example of convergent
evolution in spiders — specific differences
are found even in closely similar webs of
different species and different families
within species.
While it appears easier to distinguish
spider species by their morphological char-
acters, their behavior, at least as measured
in the pattern of the orb web throughout
life, contains potentially highly discrimina-
ble features.
454
Witt,, Rawlings, and Reed
REFERENCES
Breed, A. L., V. D. Levine, D. B. Peakall, and P. N.
Witt. 1964. The fate of the intact orb-web of the
spider Araneus diadematus Cl. Behaviour
23:43-60.
Christiansen, A., R. Baum, and P. N. Witt. 1962.
Changes in spider webs brought about by mes-
caline, psilocybin and an increase in body weight.
J. Pharmacol. 136:31-37.
Eberhard, W. G. 1971. Senile web patterns iti Ulo-
borus diversus. Develop. Psychobioll’T:245-254.
Rawlings, J. O., and P. N. Witt. 1972. Preliminary
data on a possible genetic component in web-
building. Colorado Symposium on Behavior Gen-
etics: Simple Systems. Colorado Associated Uni-
versity Press. (In pressera
Reed, C. F., P. N. Witt, and M. B. Scar boro. 1969.
The orb-web during the life of Argiope aurantia
(Lucas). Develop. Psychobiol. 2:120-129.
Reed, C. F., P. N. Witt, M. B. Scarboro, and D. B.
Peakall. 1970. Experience and the orb-web. De-
velop. Psychobiol. 3:251-265.
Wiehle, H. 1927. Beitraege zur Kenntnis des Rad-
netzbaues der Epeiriden, Tetragnathiden und
Uloboriden. Z. Morphol. Oekol. Tiere 8:468-537.
Witt, P. N. 1956. Die Wirkung von Substanzen auf
denKfetzbau der Spinne als biologischer Test.
Springer Verlag, Berlin.
WitH P. |1| 1971. Instructions for working with
web-building spiders in the laboratory. Bio-
Science 21:23-25.
Wit||p. N., and R. Baum. 1960. Changes in orb
webs of spiders during growth (Araneus diade-
micdus Cl. and Neoscona vei^braPa McCook) .
Behaviour 16:309-318.
Witt, P. ®, and C. F. Reed. 1965. Spider-web
building. Science 149:1190-1197.
WitSp. N., C. F. Reed, and D. B. Peakall. 1968. A
spider’s web. Springer Verlag, Berlin.