Images Collection


View this article in Search Friendly Plain Text
NOTE: This plain text article interpretation has been digitally created by OCR software to estimate the article text, to help both users and search engines find relevant article content. To read the actual article text, view or download the PDF above.
Reprinted from The Journal of Experimental Zoology Yol. 156, No. 3, August 1964
Composition, Function and Glandular Origin of the Silk Fibroins of the Spider Araneus diadematus Cl.1
DAVID B. PEAKALL 2
Department of Pharmacology, State University of New York, Upstate Medical Center, Syracuse, New York
The aim of this paper is to correlate the function and histology of the silk glands of the spider, Araneus diadematus Cl, with the amino acid composition of the silk. This work was done in order to place on a firmer basis the work on the effects of drugs on the rate of synthesis of protein in the silk glands of spiders (Peakall, ?64).
There is a good deal of information on the anatomy and position of the silk glands. The shape and position of the silk glands of a spider of the genus Araneus are given in many texts. All appear to be based largely on the work of Apstein (1889) and contain few original observations. Apstein noted five histologically distinct sets of silk glands in the adult female spider. There are aciniform, aggregate, ampullate, cylindrical and pyriform glands.
Specific purposes for silk from each set of glands have been proposed, (Comstock, ’48 ) although the evidence on which this is based is circumstantial. It appears to be based on observation of which spinneret is used combined with a knowledge of the gross anatomy of the silk glands. In this paper serial sectioning after a specific silk has been used permits definite correlation of glands with type of fibroin.
The amino acid composition of the total web, egg cocoon and safety thread of Ara-beus diadematus has been measured by Fischer and Brander (’60). Data on the amino acid composition of fibroins from many species of spider have been published by Lucas, Shaw and Smith (’60). They used silk from normal cocoons, egg cocoon, communal cocoons and reeled material of unknown functional form. In the present study analysis was made of threads of well-defined origin and function of a single species, Araneus diadematus.
MATERIALS AND METHODS
(1) Collection of spiders and thread
The spiders, adult female Araneus diadematus Cl, were caught locally. They were housed in cages, 20″ X 20″ X 4″, consisting of fine aluminum mesh sides and removable glass doors (20″ X 20″) front and back. These cages give the spider enough room for web-building and it is possible to open the cages without damaging the web. The spiders were fed on sugar water (10 ulofa 10% solution) and flies raised in the laboratory. On this diet they could be kept in good condition for many months.
Material was collected for examination of fibroin composition and condition of the glands at various stages. These were as follows:
(1) After completion of the web.
S||2) After completion of the scaffolding of the web.
(3) Thread pulled directly from the spider.
(4) Swathing thread.
(5) Frame threads collected from completed webs.
Ill 6) Thread used for the egg-sac.
In all but the last two cases spiders were killed for sectioning immediately after the collection of the fibroin. The six categories will now be considered in more detail.
The webs were built in the morning at first light and in summer most of the spiders built daily. Both webs and spiders were taken within a few minutes of the completion of the web. The sequence of actions involved in the formation of an
1 This investigation was supported in part by research grant PN-5234 from the National Institutes of Health, U. S. Public Health Service.
2 Supported by USPHS Institutional Grant.
345
J. Exp. Zool., 156: 345-352.
346
DAVID B. PEAKALL

Figure 1
a The appearance of the web just after completion of the scaffolding. The first three threads of the viscid spiral (indicated by the arrow) have been laid down. Material for examination of the scaffolding is taken before any viscid thread is used.
b Completed web with names of parts as used in the text.
orb web have been described in detail (Savory, -52). The important point so far as this work is concerned is that the spider pauses after the completion of the dry spiral. Then the spider completes the web by laying down the viscid spiral. It is at the stage between the completion of the scaffolding and the starting of the viscid spiral that the webs and spiders were collected for category 2. Figure la illustrates the condition of the web at the start of the laying down of the viscid spiral.
Thread for category 3 was obtained by pulling directly from the spider by attaching the end of the thread to a glass rod which was turned by a small motor. The spider was held by hand until all the thread had been rolled onto the glass rod. The spiders were killed for sectioning immediately after pulling. Only spiders that had not built during the previous 24 hours, thus presumably having full glands, were used in this experiment.
The swathing silk is that fibroin which is used to encase prey. A fly was given to a spider, after the fly had been encased it was removed from the cage and the thread carefully taken off the fly for analysis. The process was repeated until the spider refused to swath the fly. Then the spider was killed for sectioning.
The frame thread was obtained by cutting the support thread at the outermost spiral and rolling the thread onto a glass rod back to the edge of the cage. Figure lb shows the position of this thread. It was not possible to obtain a spider for sectioning just after this thread had been built because frame and radii are built together.
The thread immediately around the egg-sac was cut away from the egg-sac itself. No spider was observed during this process and thus no spider was available for sectioning.

SILK FIBROINS OF ARANEUS DIADEMATUS
347
(2) Amino acid composition of fibroins
The various fibroins, obtained as described in the previous section, were collected several times from the same spider. Three to five samples were pooled for each determination. Thus it was possible to compare the composition of the various fibroins from the same spider. The determinations were repeated on five different spiders.
The fibroins were hydrolysed in boiling 6N hydrochloric acid for 18 hours. The solution was then evaporated just to dryness and the amino acids dissolved in 10% aqueous n-propyl alcohol. The examination for the presence of tryptophan was made after hydrolysis with 14% barium hydroxide.
The amino acid composition was determined by ascending paper chromatography using a system essentially similar to that of Mitzell and Simpson (’61). Two minor changes were made, the ratios of the components of the first solvent were altered to give better resolution, and the substitution of ammonia for cyclohexamine vapor to improve the clarity and permanance of the dyeing. Whatman no. 1 paper was used and the material was applied by hand using a micropipette to a circle 3 mm in diameter. The material was contained in 6 nl of solvent. The solvent systems used were as follows: — first nBuOH: HAc: H20 = 120:30:50 and second nBuOH: EtMeCO: H20: NH3 = 80:80:40:1. The paper was dried at room temperature in a forced draft after each solvent system. At the completion of the run the paper was dipped in 2.5% solution of ninhydrin, dried at room temperature and then heated for three minutes at 60°.
The optical density of the spots was measured 24 hours later using a photovolt transmission densitometer. The area under each peak recorded by this instrument was measured with a planimeter and compared with standard spots run in the same tank. The results were corrected to zero hydrolysis time. Six to ten duplicate runs were made and the variation for the major components found to be ± 5%. The method used in calculating the amino acid composition from this data was that of Fisher, Parsons and Holmes (’49) and the
validity of this procedure has been considered in detail by Block, Durrum and Zweig (’58).
(3) Histological
The spiders were fixed in buffered 10% formaldehyde for 7 to 10 days. The abdomens were double embedded using collodion and paraffin and then serial sectioned at 6 n. Alternate slides were stained with azure B and hematoxylin and eosin. These serial sections were then examined to determine the general configuration of the silk glands. Knowing which fibroins had been used immediately before death it was possible to correlate the fibroin with the gland since the amount of protein in the specific gland used would be depleted.
During the course of preparation of sections the protein contained within the glands shrinks and leaves an empty area along the walls of the glands. This artifact can be seen in figure 2. It is observed that the protein platelets frequently fall out of the section. By examining several consecutive sections, however, it has been established which glands are empty and which are full. As a consequence of this loss of protein platelets individual sections (such as those shown in figure 2a—d) often show more empty glands than really exist.
RESULTS (1) Amino acid composition
The amino acid composition of the various fibroins is given in table 1. It will be noted that the composition of the frame thread, pulled thread and the material used in the web up to the completing of the scaffolding are essentially identical. These threads are characterized by high values of the three principal amino acids, alanine, glycine and glutamic acid. The total web varies from the scaffolding threads in having a much lower value for glutamic acid and a much higher value for proline and isoleucine. The egg cocoon fibroin has high serine and low glycine content as compared to the web proteins. The swathing silk also has a higher serine and lower glycine content than the web proteins but not to such a marked extent as the cocoon proteins.
348
DAVID B. PEAKALL

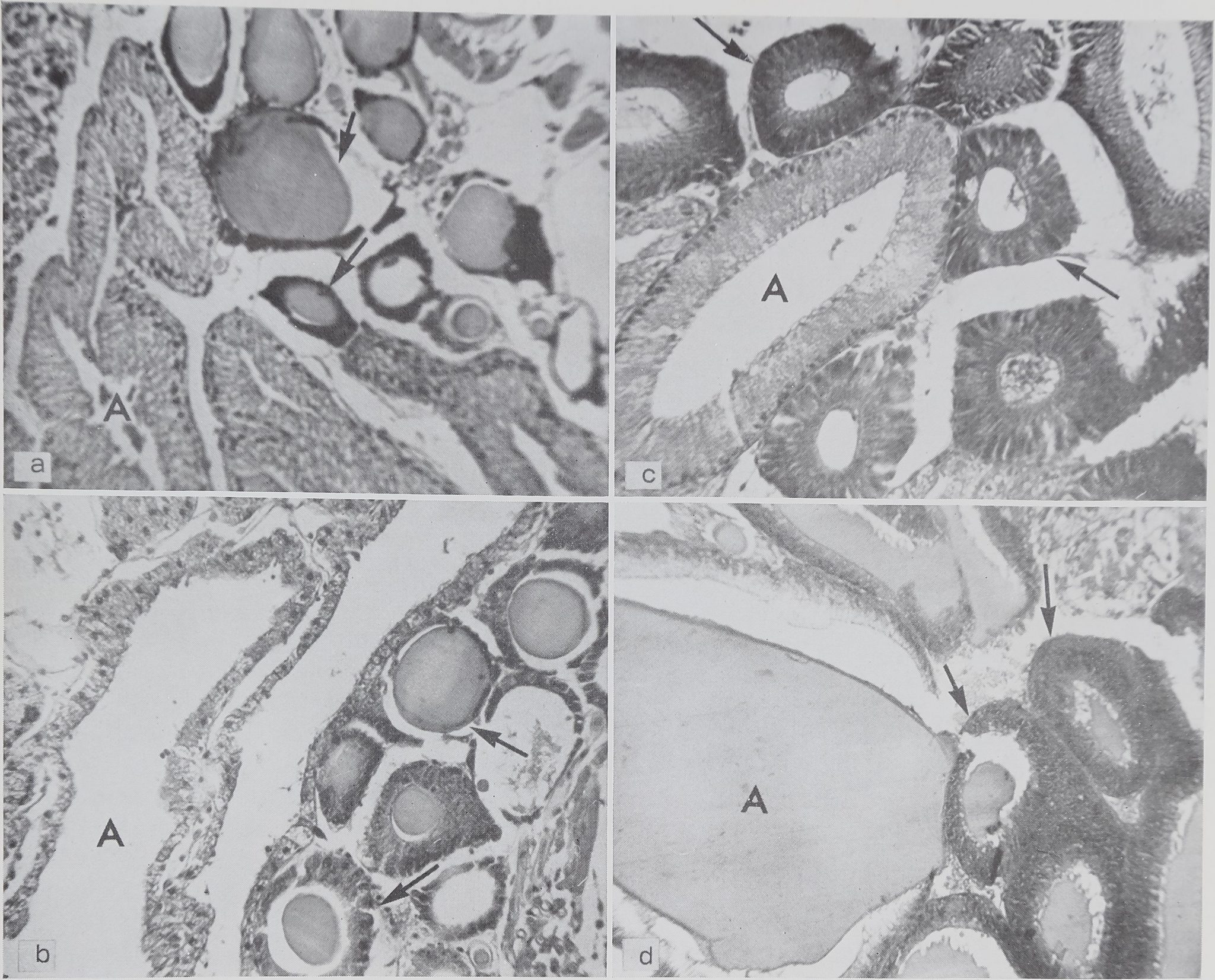
Fig. I Sections showing the condition of both ampullate and aggregate, glands after various fibroins ha* been used. Ampullat^^*|ids are labeled “A” and aggregate glands are indicated by arrows. Staining hematoxylin and eosin. Magnification 700
a From spider killed immediately after completing the scaffolding of the web. The ampullate glands (A) are empty and the aggregate glands (arrows) are full.
b From spider killed after emptying the glands by pulling the thread. Condition of glands as (a), c From spider killed immediately after doirfpleting the web. Both ampullate (A) and aggregate (arrows) glands are empty.
d From spider killed after a period when no silk was used. Both ampullate (A) and aggregate (arrows) glands are full.
(2) Histological
The technique of making serial sections of the spider’s abdomen after a specific silk has been used gives a clearer picture of the histology of the glands than has been possible by observation and gross dissection. Examination of these serial sections shows three points that add to previous knowledge of the subject.
(1) Comparison of sections from spiders killed after completing the scaffolding of the web and after completing the whole
web clearly show that the viscid thread is produced wholly or in part in the aggregate glands. This is shown in figures 2a and 2c.
(2) Comparison of sections of spiders killed after completion of the scaffolding of the web and after the thread had been removed by pulling shows that, in both cases, the thread came from the ampullate glands (figs. 2a and 2b).
(3) The swathing silk glands were found to be different from those previously described for the genus Araneus. The spin-
SILK FIBROINS OF ARANEUS DIADEMATUS
349
TABLE 1
Amino acid composition in g/100 g of various silk fibroins of the spider, Araneus diadematus Cl.
Total Frame Pulled Scaffolding Egg
web thread thread of web cocoon
| Alanine | 27.3 | 32.8 | 32.7 | 33.4 | 25.4 | 25.3 |
| Arginine | 3.4 | 4.2 | 3.2 | 3.6 | 5.0 | 3.8 |
| Aspartic acid | 1.8 | 1.3 | 1.1 | 1.3 | 2.3 | 2.1 1 A I |
| Glutamic acid | 9.1 1 | 17.7 | 17.9 | 17.8 | 13.6 1 | 11.0 1 |
| Glycine | 20.1 | 23.8 | 24.3 | 24.3 | 11.9 1 | 15.3 1
A O 1 |
| Isoleucine | 5.7 1 | 2.1 | 1.7 | 2.4 | 5.0 1 | 4.8 1 |
| Leucine | 2.9 | 1.8 | 2.1 | 1.5 | 3.5 | 3.2 |
| Lysine | 3.2 | 2.1 | 1.8 | 1.9 | 1.3 | 2.2
n.d. |
| Phenylalanine | 1.1 | n.d. | n.d. | n.d. | n.d. | |
| Proline | 12.8 1 | 3.1 | 3.1 | 2.3 | 3.8 | 10.2 1 |
| Serine | 5.3 | 5.8 | 6.3 | 6.4 | 18.7 1 | 11.7 1 |
| Threonine | 2.1 | 1.8 | 2.1 | 1.7 | 3.1 1 | 1.9 |
| Tyrosine | 1.7 | 2.4 | 1.8 | 2.1 | 3.7 1 | 3.7 1 |
| Valine | 3.|U | 1.1 | 1.9 | 1.3 | 2.7 | 4.8 1 |
| PQFigures considered thread. | to deviate significantly from the frame | thread, pulled thread and scaffolding | ||||

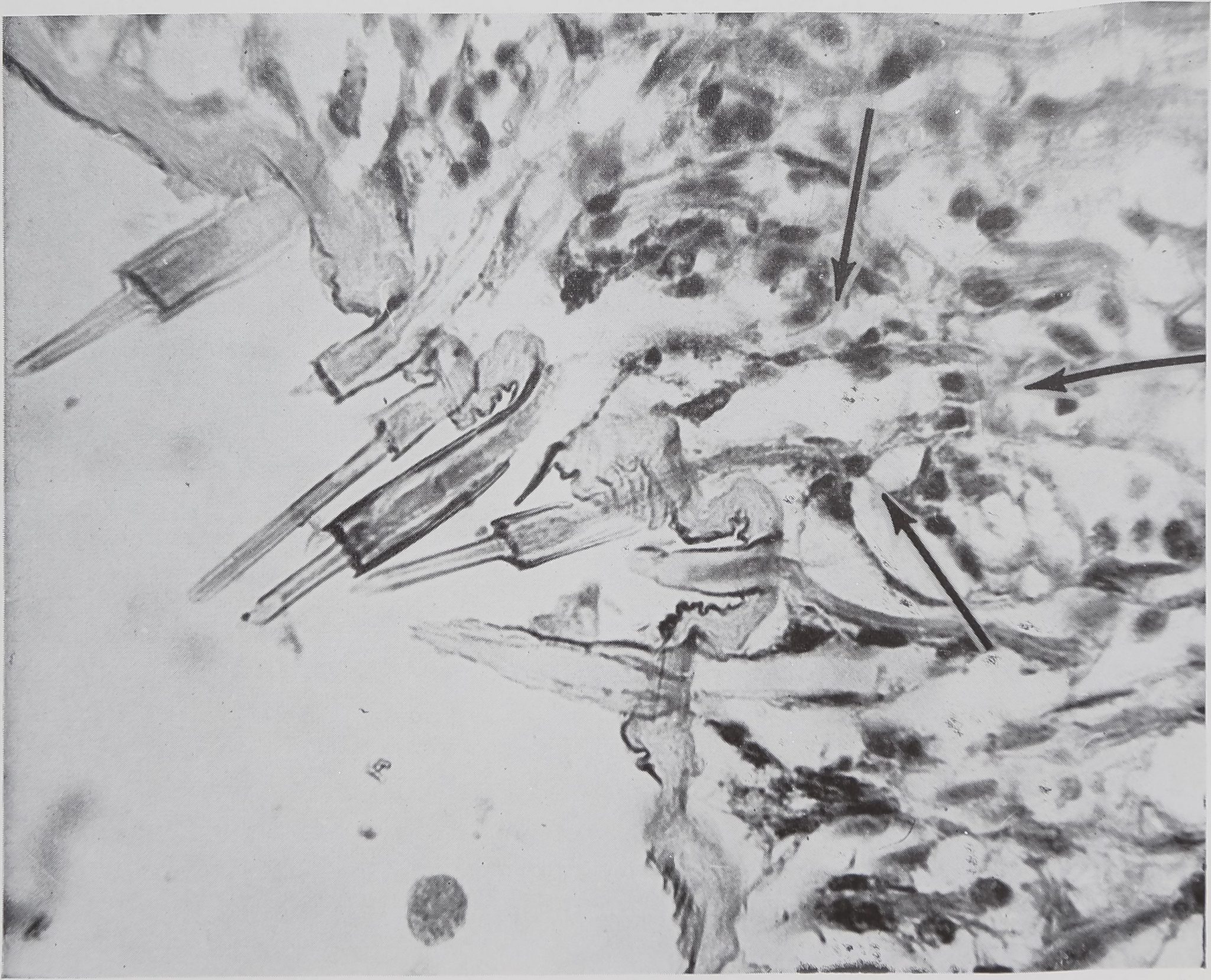
Fig. 3 Structure of empty swathing silk glands. These are short glands, lying in clusters close to the wall of the spinneret. Each spinning tube, several of which can be seen in the photograph, is connected by a short, unbranched duct to the lumen of the gland. The outline of one of these glands is indicated by the arrows. Stain, Azure B. Magnification 350 X.
ning tubes are type “a” (using the classification of Comstock, ’48) and are found on the hind and middle spinnerets as has been noted by other observers. However the aciniform glands with multiple sacs
emptying into a common duct at some distance from the spinneret as described by Comstock (’48) for the family Aranea were not observed. Rather, as is illustrated in figure 3, each spinning tube has a single
DAVID B. PEAKALL
350
small gland which lies just inside the wall of the spinneret. That these glands are the site of the swathing silk has been demonstrated by the fact that these glands, normally full of protein, are empty after the spider has swathed several flies.
Two other types of glands noted by previous investigators were seen but not studied. These were the cylindral glands which are considered responsible for the cocoon silk and the small pyriform glands which are considered to make silk for the attachment discs.
DISCUSSION (1) Amino acid composition
The amino acid composition determinations given in table 1 for the total web, frame and egg cocoon threads agree fairly well with those of Fischer and Brander (’60). The only serious difference being the low proline value found for the frame thread compared to the total web which was not found by Fischer and Brander. The amino acid composition of pulled thread, the scaffolding of the web and swathing silk had not previously been determined.
The similar amino acid composition of the pulled thread, frame thread and scaffolding of the web support the view that these materials are identical. The variation from spider to spider for the various fibroin compositions was within the experimental error of The amino acid composition of the other fibroins shows that these are all quite different proteins; the differences in the paper chromatograms could be readily seen by eye. However, the typical silk pattern of high alanine and glycine is found in all of them. It is possible that each of these fibroins is, in fact, a group of proteins rather than a single protein. To test this the electrophoretic separation of material from the ampullate gland was attempted and only a single, although somewhat diffuse, peak was found. War-wicker (private communication), on the basis of x-ray diffraction studies, considers that each gland produces a single protein.
Any work on protein taxonomy of the fibroins of spiders must clearly take into account the type of fibroin used. This is illustrated by the wide differences found by Lucas, Shaw and Smith (’60) between
Nephila madgascariensis and Nephila sene galensis. The fibroin of the former was supplied reeled and probably corresponds to pulled thread whereas the fibroin of the latter was from the egg cocoon. The differences were, therefore, probably not one of species but rather of thread type. The most convenient protein for taxonomic work would be the pulled thread. It can be obtained when required and is free from contamination that can accompany webs and cocoons and thereby removes the necessity for cumbersome and undesire-able cleaning processes.
(2) Histological
Comparison of serial sections of spiders killed after the glands had been emptied by pulling and after completion of the scaffolding of the web shows that the material has come from the ampullate glands in both cases (fig. 2a and 2b). It was found that the ampullate glands were not completely emptied at the stage of completion of the scaffolding. No quantitative measurements were made but examination of serial sections of three spiders collected at this stage gave the impression that two-thirds to three-fourths of the material had been used. Koenig (’51) allowed spiders to build webs to the stage where all but the last three radii had been laid down. Then he continuously eliminated the last three radii as soon as the spider had built them. He found that the spider would persist in laying down radii until it had built two and a half times the normal number of radii. Since the web at the stage when the scaffolding is completed consists of the frame threads, the temporary spiral, the hub and radii (see fig. la and b), the radii probably represent no more than 50% of the material used at this stage. Thus Koenig’s observations are in reasonable agreement with the histological data presented here.
It has been suggested (Comstock, ’48; Peters, ’55) that the aggregate glands are responsible for the viscid thread because these glands occur only in the orb-web builders. Comparison of sections of spiders killed after the completion of the scaffold-
SILK FIBROINS OF ARANEUS DIADEMATUS
ing with those killed after the completion of the total web confirms this hypothesis. The aggregate glands are found to be full in the case of the spider killed after completion of the scaffolding of the web (fig. 2a) and largely empty after completion of the web (fig. 2c). The apparently empty aggregate glands in figure 2a are due to loss of protein platelets during preparation (see methods).
This still leaves open the question of whether the viscid spiral consists of two components, a central support thread from the ampullate gland coated with a viscid substance from the aggregate gland, or a single sticky thread. Nephila Peters (’55) considered that the former was the case. He found that the structural thread came from a special gland called the cor-onatae gland and the glue from the aggregate gland. Richter (’56) has presented some photographic evidence for the exist-ance of three layers, a structural thread, a viscid layer and an outer muscous layer. However, the value of this evidence cannot be assessed since no experimental details are given nor is the name of species of spider stated. Working with serial sections of Araneus we found no evidence for the existence of a coronatae gland. If this is correct then the sticky spiral could be made of material from the ampullate glands covered with material from the aggregate glands, or simply of material from the aggregate glands. Koenig (’51) found that the sticky spiral was very small after the radii efimination experiment. It could be inferred that this is due to the shortage material in the ampullate glands. However, since the experiment interfered with normal web-building patterns, this is not conclusive evidence. Examination of sections of spiders taken after the normal completion of the scaffolding and after completion of the web indicates that less material remains after completion of the web than at the stage of completion of the scaffolding. The point has not been proved conclusively, but our observations support the view that the viscid spiral consists of material from the ampullate glands coated with material from the aggregate glands.
SUMMARY
1. The amino acid composition of various fibroins of the spider Araneus diadem-atus Cl were determined. It was found that the thread pulled from the spider had an identical composition with that of the scaffolding of the web. However the composition of the scaffolding was different from that of the complete web, showing that at least two distinct proteins are used in the formation of the web.
2. The glands responsible for each fibroin were located histologically, by serial sectioning, after a certain fibroin had been depleted. It was found that the pulled thread and the material used for the scaffolding of the web both came from the ampullate glands. The postulate that the viscid thread comes wholly or in part from the aggregate glands was confirmed. It seems likely that the sticky spiral consists of thread from the ampullate glands coated with material from the aggregate glands.
3. The structure of the swathing silk glands was found to be different from that previously described.
ACKNOWLEDGMENT I should like to thank Dr. Peter Witt for his help and encouragement throughout the course of this study.
LITERATURE CITED
Apstein, C. 1889 Bau und funktion der Spinndrüsen der Araneida. Arch. Naturg., Berlin, 55: 29-74.
Block, R. J., E. L. Durrum and G. Zweig 1958 A manual of paper chromatography and paper electrophoresis. Academic Press. New York. Comstock, J. H. 1948 The Spider Book. Comstock Publishing Co. Ithaca.
Fischer, F. G., and J. Brander 1960 Eine Analyse der Gespinste der Kreuzspinne. Z. Physiol. Chemie., 320: 92-102.
Fisher, R. B., D. S. Parsons and R. Holmes 1949 Quantitative paper chromatography. Nature, 164: 183.
Lucas, F., J. T. B. Shaw and S. G. Smith 1960 Comparative studies of fibroins. 1. The amino acid composition of various fibroins and its significance in relation to their crystal structure and taxonomy. J. Mol. Biol., 2: 339-349. Mitzell, M., and S. B. Simpson, Jr. 1961 Paper chromatographic separation of amino acids, A solvent to replace phenol. J. Chromatog., 5: 157-160.
Peakall, D. B. 1964 Effects of cholinergic and anticholinergic drugs on the synthesis of silk
352
DAVID B. PEAKALL
ñbroins of spiders. Comp. Biochem. Physiol., in press.
Peters, H. M. 1955 Uber den Spinnapparat von Nephila madagascariensis (Radnetzspinnen, Fam. Argiopidae). Z. Naturforsch., 10b: 395-404.
Richter, G. 1956 Untersuchungen über Struktur und Funktion der Klebefäden in den Fanggeweben ecribellater Radnetzspinnen. Natur-wiss., 43: 23.
Savory, T. H. 1952 The Spider’s Web. Warne. London.