NOTE: This plain text article interpretation has been digitally created by OCR software to estimate the article text, to help both users and search engines find relevant article content. To read the actual article text, view or download the PDF above.
Rev. Comp. Animal — T. 5 — Ier Trim. 1971 — pp. 19 à 26 — Crépln-Leblond et Cie, Edit. Paris
CONSEQUENCES HISTOLOGIQUES ET COMPORTEMENTALES
DE LESIONS (par laser) DU GANGLION SUS-OESOPHAGIEN
DE L’ARAIGNEE (ARANEUS DIADEMATUS CL)
LE OUELTE (L.), et WITT (P,N.)
Laboratoire de Psychophysiologie,
Faculté des Sciences, Nancy
Résumé
Trente six araignées femelles adultes (Araneus
diadematus) qui avaient construit, dans les con-
ditions constantes du laboratoire, au moins qua-
tre toiles orbiculaires, ont été soumises à l’im-
pact d’un faisceau laser d’énergie, de longueur
d’onde, et de diamètre connus. L’analyse histo-
logique du céphalothorax révèle des lésions pro-
fondes et circonscrites dans le ganglion sus-
oesophagien des 12 animaux. Ces mêmes sujets
manifestent des troubles graves et persistants
dans leur comportement, et on peut les me-
surer d’après les changements importants de
la taille et de la régularité des toiles qu’ils filent
ensuite. Les auteurs croient que les corrélations
entre l’analyse histologique des lésions causées
par le laser et l’évaluation des mensurations de
la toile avant et après ces lésions chez Araneus
fournissent un moyen d’étudier les rapports en-
tre les structures et les fonctions dans le sys-
tème nerveux des invertébrés.
Summary
Thirty-six adult female spiders (Araneus diade-
matus Cl) which had built under constant labo-
ratory conditions at least four orb webs, were
subjected to the impact of a laser beam of known
energy, wave length, and focal size.
Histological analysis of the céphalothorax re-
vealed deep, circumscribed lesions in the supra-
esophageal ganglion of 12 animals. The same ani-
mals manifested severe and persistent behavioral
disturbances afterwards, wich could be measured
as significant changes in size and regularity of
post-laser webs. The authors believe that corre-
lation of histological analysis of laser damage
with evaluation of web measurements before and
after lesions in Araneus provides a means of
studying structure-function relationships in the
nervous system of invertebrates.
Zusammenfassung
36 adulte Spinnenweibchen (Araneus diadema-
tus Cl), die unter konstamen Laboratoriumsbe-
dingungen mindestens vier Netze gebaut hatten,
wurden der Wirkung eines Laserstrahls mit be-
kannter Energie, Wellenlànge und Brennweite
ausgesetzt.
Bei der histologischen Untersuchung des Cépha-
lothorax konnten tiefe, ortlich begrenzte Lasionen
im supraoesophagen Ganglion von 12 Tieren fest-
gestellt werden. Diese Tie re zeigten starke, anhal-
tende Verhaltensstôrungen, die in signifikanlen
Verànderungen der Grosse und Regelmassigkeit
der Netze nach der Laserbehandlung zum Aus-
druck kam. Die Autoren sind der Ansicht, dass
die Untersuchung der Beziehung zwischen den
histologisch feststellbaren Laserbeschàdigungen
und den Netzmassen vor und nach den Lasionen
bei Araneus ein Mittel zur Erforschung der Strut-
tur-Funktionsbeziehungen im Nervensystem der
Invertebraten darstellt,
19
Introduction (1)
Dans l’étude des conséquences de lésions
du système nerveux central (SNC), il est
rare que la lésion et le comportement
soient en même temps évalués qualitati-
vement et quantitativement. Le problem
me, au point de vue histologique, peut ré-
sider dans la difficulté de mesurer exac-
tement l’étendue d’une destruction des
tissus. Il réside également dans le fait que
la cible est parfois une structure plus ou
moins polymorphe de grandes dimensions
et qui peut être seulement évaluée gros-
sièrement. Le comportement est souvent
très facilement supprimé après lésion. Il
peut ne pas être suffisamment rigide
pour permettre une analyse quantitative
ou être trop simple pour permettre une
évaluation de la nature des divers désor-
dres occasionnés par les lésions. L’Arai-
gnée construisant une toile géométrique
possède un système nerveux hautement
centralisé qui comprend un nombre rela-
tivement faible de cellules si on le com-
pare au système nerveux des Vertébrés.
Des recherches antérieures sur l’anatomie
et l’histologie du système nerveux central
des Arachnides par Babu (1965), Meier
(1967) et Le Guelte et Witt (1968) nous
ont fourni les connaissances nécessaires
sur la structure « normale )> du jjferveau
de l’Araignée.
Le laser a été employé chez d’autres
Invertébrés, en particulier sur des Crus-
tacés (Balesdent, 1968), cependant, aucun
travail de ce genre n’a été réalisé sur
l’Araignée, matériel particulièrement dif-
ficile à opérer (Legendre, 1958). Ces tra-
vaux et ceux d’autres auteurs, ont montré
que l’énergie importante fournie par le
rayon laser est un outil utile pour la mi-
crochirurgie chez les Invertébrés, mais
rien ne permet de prévoir, à ce jour, la
nature et l’étendue possible d’une lésion
chez Araneus diadematus. Notre travail se
présente donc comme une première tenta-
tive pour établir si la chirurgie à l’aide •
du laser est adéquate pour notre animal
et nous souhaitons qu’il ouvrira la voie
pour une microchirurgie plus fine.
La structure des toiles et leurs variations
utilisées comme indications de l’aptitude
de l’animal normal à s’adapter à des trau-
matismes chimiques ou mécaniques, et
l’évaluation de la plasticité du comporte-
ment, ont été l’objet de recherches dans
les laboratoires des deux auteurs depuis
plusieurs^ années. On a pu ainsi montrer
que, même chez des animaux qui sont gra-
vement handicapés, un certain type de
construction est mis en œuvre, et que des
toiles mesurables sont construites. La
construction de la toile est une nécessité
si impérative pour la survie de l’animal
qu’elle peut être difficilemenu éliminée.
Elle est aussi suffisamment complexe pour
permettre une différenciation entre des
changements de différentes natures : l’es-
pacement régulier des rayons, par exem-
ple, peut être modifié sans qu’il y ait de
variations dans la régularité des spires et
les dimensions. D’autre part, la taille de
la toile peut être modifiée, par une subs-
tance tranquillisante par exemple, sans
changement dans sa régularité.
Ces considérations nous ont amené à
penser que les lésions, au laser, du S.N.C.
de l’Araignée, pourrait fournir un bon mo-
dèle pour l’étude des interrelations entre
structure nerveuse et fonction ; cet article
est une étape dans une série où les au-
teurs utiliseront l’Araignée comme mo-
dèle pour l’étude du comportement et de
son codage.
(I) Les travaux rapportés ici ont été effectués
dans le laboratoire du Dr Witt, Department of
Mental Health, Division of Research, Raleigh,
N.C. — U.S.A.
Materiel
Les lésions ont été localisées sur 36
Araignées? parmi celles qui tissèrent un
20
ce sont toutes des femelles adultes. 14 se
sont révélées avoir des lésions au niveau
du ganglion sus-ce sophagien (position C3,
voir >Le Guelte et Witt, 1968). Deux
de cel’les-ci n’ont .montré aucun change-
ment dans la toile après la lésion qui était
très superficielle. Les 12 autres sont étu-
diées ici.
Le laser utilisé est un laser à rubis de
6 934 angstroms, avec une énergie de 0,8
à 1 joule. Il était focalisé sur la chitine à
l’aide d’un microscope puis la mise au
point était descendue de 0,5 mm (Witt,
1969). Pendant l’opération, l’Araignée
était maintenue sur un support recouvert
de gaze, ce qui permet une visée du cé-
phalothorax. La position de l’ensemble
était réglée grâce à un micromanipula-
teur.
Méthodes histologiques
Nous avons d’abord apprécié le nombre
total de cellules nerveuses en les comptant
toutes les trois sections, l’épaisseur de
nombre de toiles suffisant avant lésion,
chaque coupe étant de 6 microns et le dia-
mètre moyen d’une cellule de 20 microns.
Dans ces conditions de mesure, le nombre
de cellules accessibles à l’aide du laser,
par la face supérieure de l’animal, est
d’environ 6 000 sur un total de 15 000
cellules environ pour l’ensemble des gan-
glions sus et sous-œsophagiens.
La recherche des lésions est faite sur
coupes histologiques colorées à l’héma-
toxyline-éosine. D’autres colorations (im-
prégnation argentique ou thionine) ne
nous ont pas apporté de meilleurs résul-
tats.
La lésion est repérée grâce à la colora-
tion plus dense des noyaux au-dessous de
la cuticule brûlée. Comme les animaux
étaient fixés deux semaines après l’opé-
ration, nous admettons que les cellules à
noyaux détruits étaient les seules à ne
plus être fonctionnelles. Leur nombre est
évalué sur les coupes par comptage au
microscope.
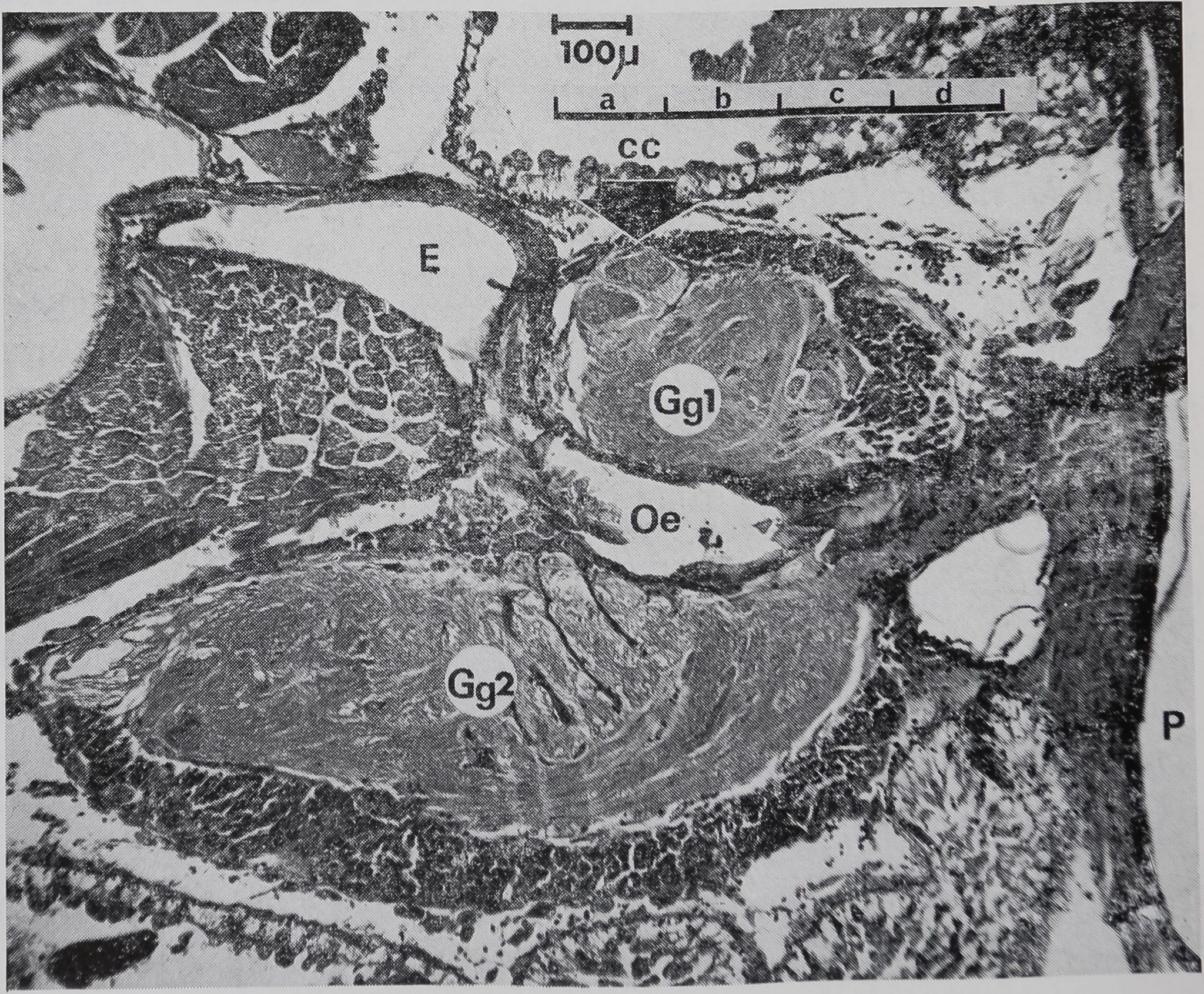
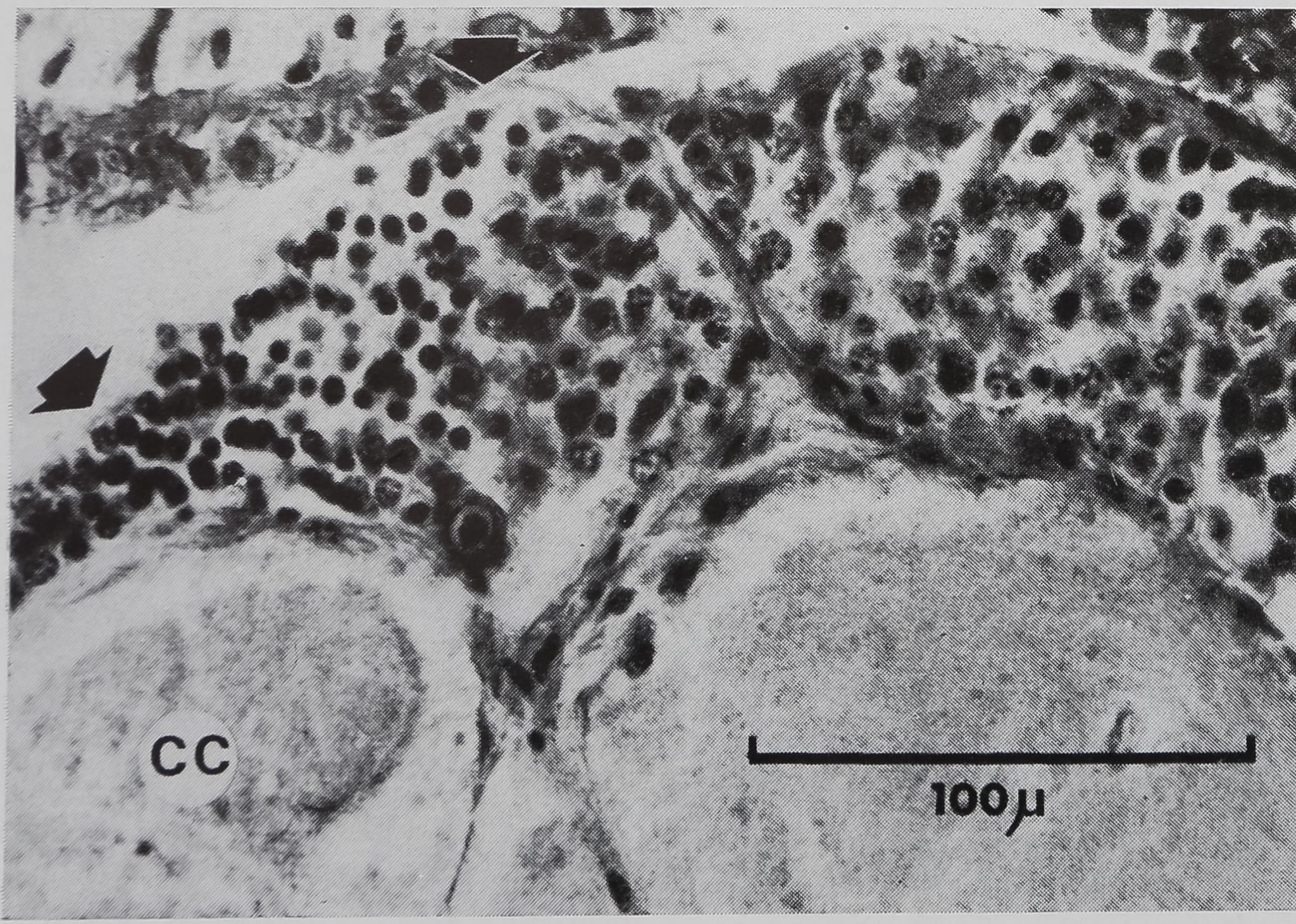
Figure 1
Système nerveux cen-
tral d} Araneus diade-
matus – Vue générale –
Coupe sagittale.
Cg 1 : ganglion sus-
œsophagien
Gg 2 : ganglion sous-
œsophagien
CC : corps central
Oe : œsophage
E : estomac
P : pharynx.
21
La figure 1 permet de localiser l’em-
placement des différentes lésions et de
noter la répartition des neurones près
du niveau de l’oesophage (coupe sagitta-
le médiane).
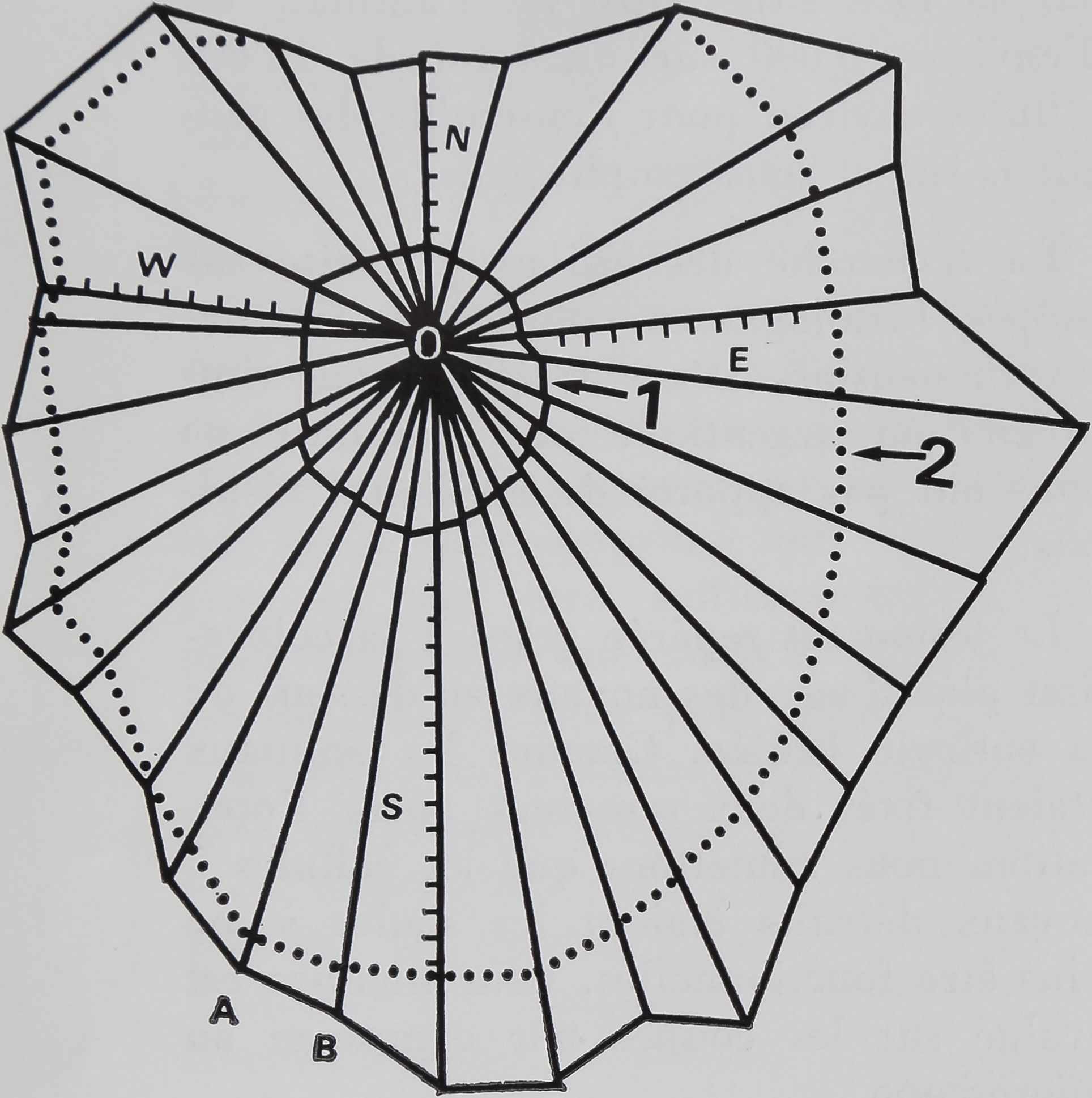
Figure 2
Mesures effectuées sur la toile en vue de
l’évaluation statistique des modifications
provoquées dans la toile par la lésion.
1 — Spirale captrice la plus interne délimi-
tant la zone libre, intérieurement.
2 — Spirale captrice la plus externe déli-
mitant la surface captrice.
N — Rayon supérieur sur l’axe vertical.
S — Rayon inférieur sur l’axe vertical. Le
rapport N/S détermine le quotient d’excen-
tricité de la toile.
W et E — Rayons horizontaux dont le rap-
port détermine le quotient d’excentricité
horizontal de la toile.
Sur les rayons N, S, W, E, la distance des
spires (en traits courts ici) par rapport au
moyeu O est mesurée et permet les calculs
de régularité des spires (Voir WITT et coll.
1968).
A O B E- Angle entre 2 rayons (AO et BO).
Méthodes d’étude du comportement
Au moins 4 toiles étaient obtenues
avant de soumettre les Araignées au la-
ser et 5 toiles après. Vingt-sept paramè-
tres étaient mesurés dans la toile et leur
comparaison avant et après lésion était
réalisée à l’aide du test T de Student et
du x2. L’ensemble des mesures était ré-
capitulé sur cartes analysées par un or-
dinateur IBM 1620-1311. La figure 2 mon-
tre comment une toile d9Araneus diade-
matus est enregistrée par l’ordinateur
(Witt et coll. 1968). Cette méthode a
permis de détecter des changements dans
la taille, la régularité, la forme et la struc-
ture fine de la toile, ceci indique quelle
fonction est perturbée chez l’Araignée
constructrice. Par exemple, une produc-
tion normale de soie et de petites toiles
observées après absorption d’une drogue
tranquillisante sont la preuve d’une mo-
dification du comportement de ‘construc-
tion. Par contre, une production réduite
de soimet de petites toiles après absorp-
tion d’atropine indique que la réduction
de la synthèse des polypeptides est le
premier impact de la drogue.
Résultats
A — Conséquences histologiques de
la lésion au laser.
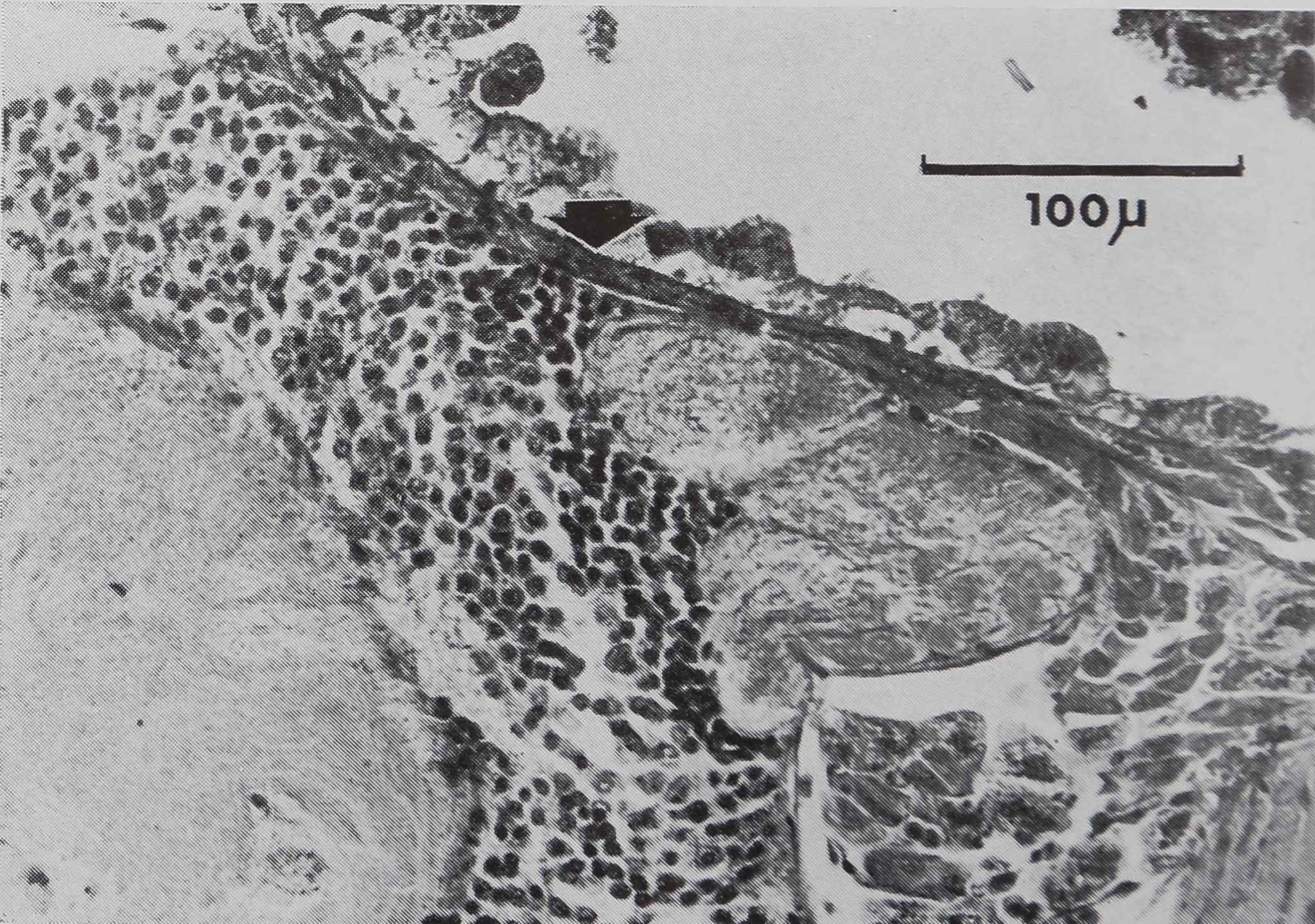
a) Traces superficielles de la lésion
Sous l’effet du rayonnement laser, la
chitine est brûlée et le rayon est réfléchi
en partie, ceci augmentant considérable-
ment la dimension de la zone modifiée.
Le eéphalothorax étant coloré par des
pigments hypodermiques, l’énergie du
rayonnement est absorbée en grande par-
tie par la chitine. Il se forme une au-
réole qui brunit progressivement au bout
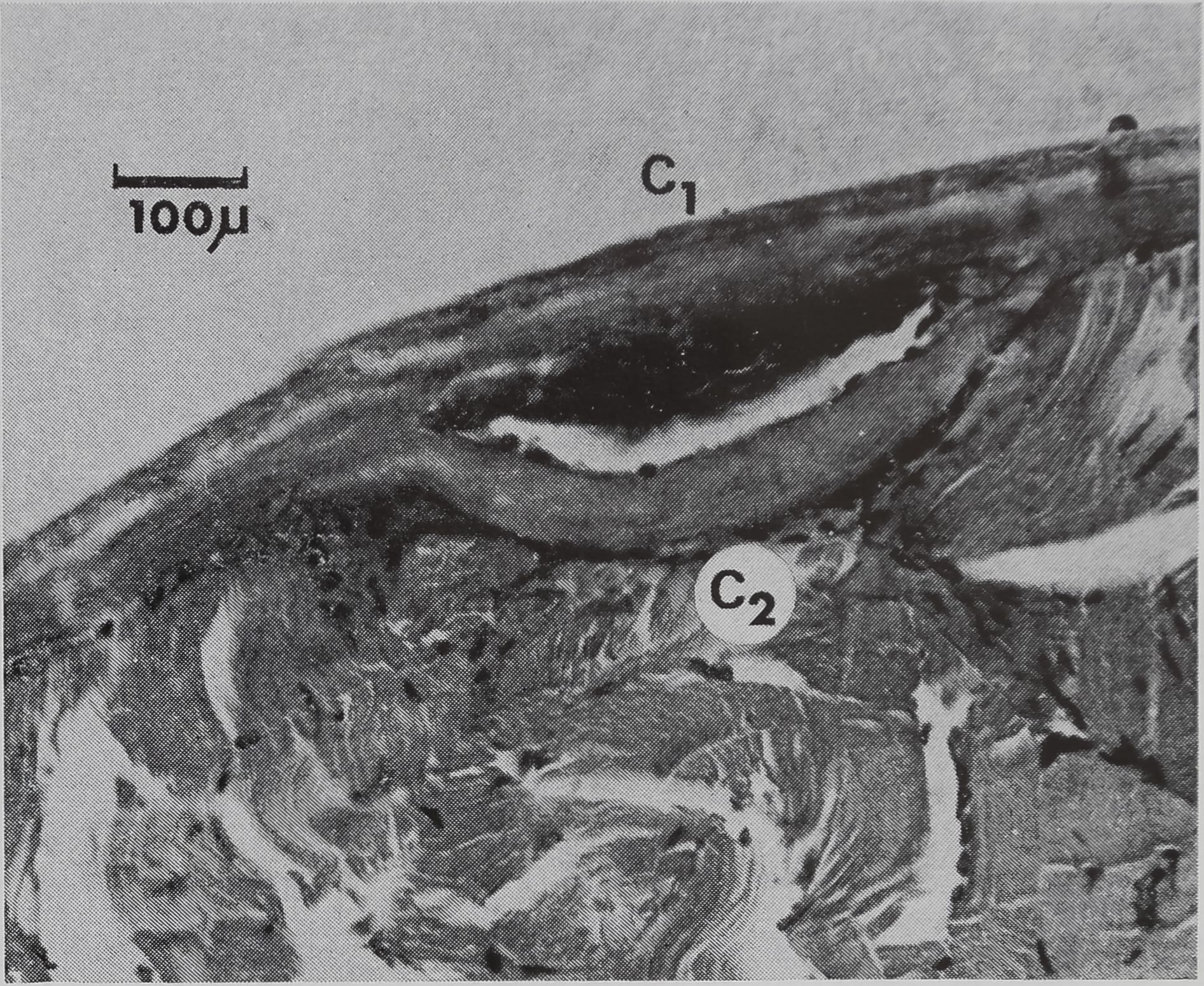
de quelques jours. Après deux semaines,
quand les animaux sont sacrifiés, on re-
marque au-dessous de la chitine nécro-
sée une zone plus sombre formée par les
tissus sous-jacents l’ancienne chitine qui
sont inclus entre l’ancienne et la nouvel-
le chitine (fig. 3). La nouvelle chitine a
été secrétée par des cellules hypodermi-
ques qui ont remplacé celles de la zone
détruite. En effet, l’hypoderme sous la
chitine brûlée, a été également détruit
comme l’indiquent les coupes pratiquées
sur des animaux fixés un ou deux jours
après l’opération.
b) Dégâts en profondeur
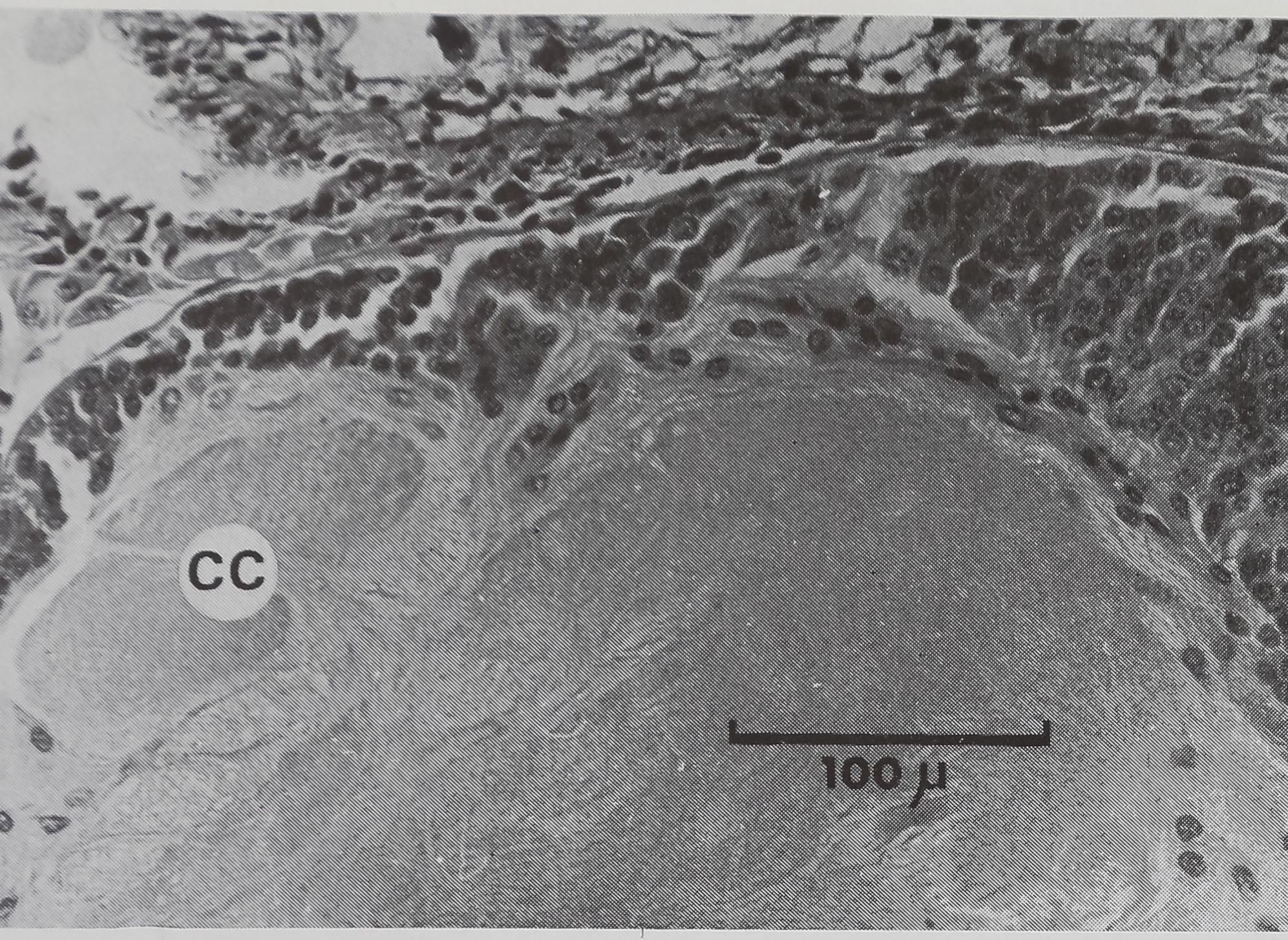
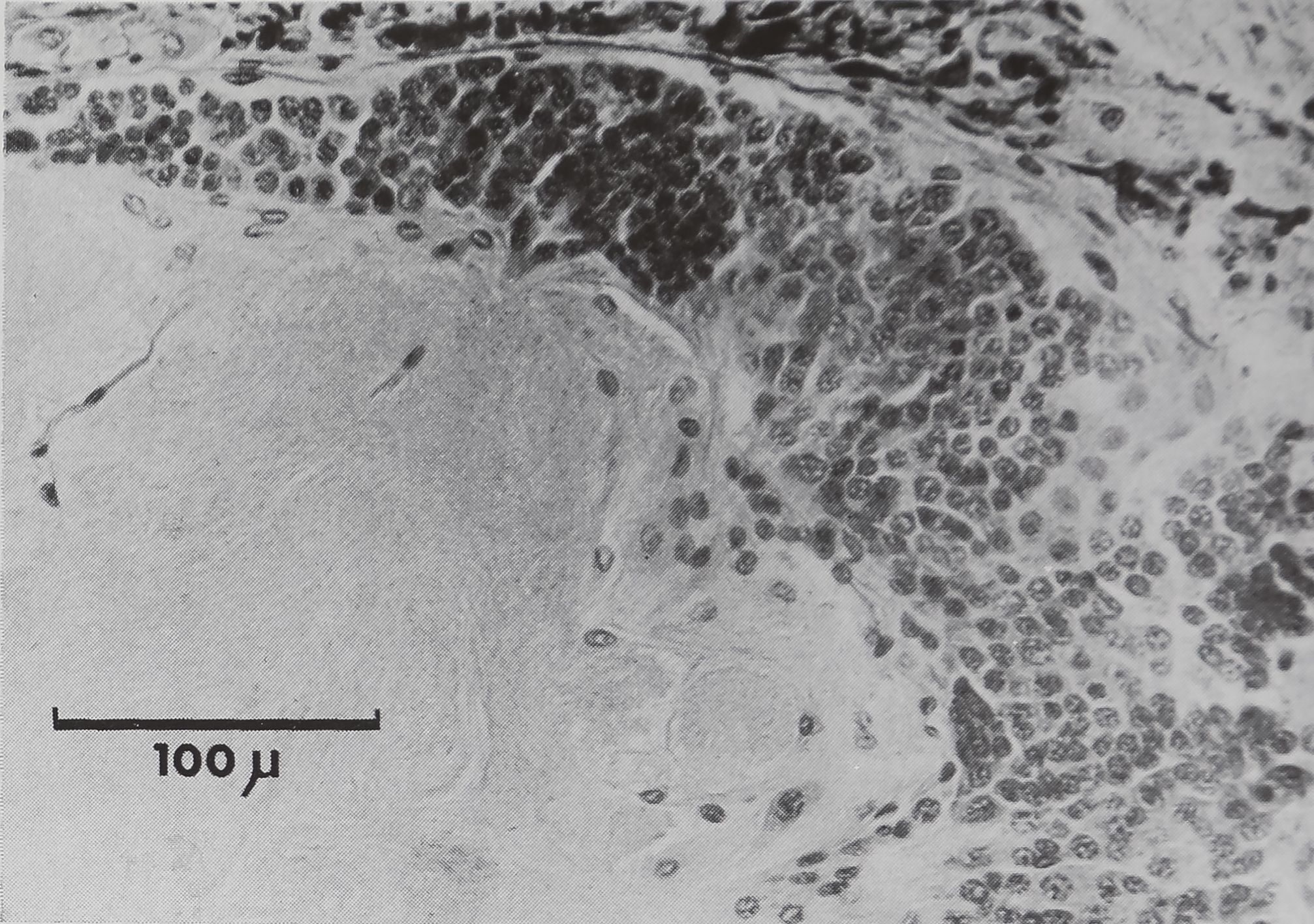
En profondeur, l’analyse histologique
permet de constater une atteinte des
noyaux de la partie supérieure du gan-
glion sus-oesophagien (comparer les fi-
gures 4 et 5 aux figures 6 et 7). La région
lésée est caractérisée par de petits noyaux
constituant une seule tache au milieu de
la cellule alors que les noyaux environ-
nants sont plus clairs et que les pigments
y sont distribués en grains moins colorés.
On doit noter qu’il existe, (à l’état nor-
mal, des cellules à peu près identiques
au point de vue taille et allure du noyau
(cellules de type A selon Babu, 1965),
mais ces cellules sont dispersées au ni-
veau du ganglion sus-œsophagien parmi
des cellules de type B de 20 microns de
diamètre dans lesquelles la chromatine
est sous forme de granulations.
Le rayon laser utilisé lors des expérien-
ces était focalisé pour avoir en surface
un diamètre d’environ 7 microns (WITT,
1969), mais, après mise au point sur la
chitine, le foyer de l’objectif était des-
cendu de 0,5 mm de sorte que la zone
nécrosée superficiellement atteint des di-
mensions de 500 microns. A la partie su-
périeure du ganglion sus-oesophagien, qui
se trouve à environ 0,75 mm au-dessous
de la chitine et qui est localisé grâce à
un quadrillage de la surface externe du
céphalothorax – Zone C 3, (Le Guelte et
Witt, 1968) la lésion a un diamètre de
80 à 100 microns.
Nous avons constaté que l’extension de
la lésion au niveau des neurones sus-
œsophagiens n’est pas liée à la puissance
du rayon laser utilisé» La traversée des
tissus sus-jacents, plus ou moins colorés,
absorbants plus ou moins l’énergie, est
responsable des différences dans la gra-
vité des lésions pour des conditions ex-
périmentales identiques.
Figure 3
Aspect superficiel d’une lésion. Coupe sa-
gittale.
C 1 : ancienne cuticule
C 2 : nouvelle cuticule.
B — Conséquences comportemen-
tales.
Toutes les araignées qui ont construit
un nombre de toiles suffisant pour l’éva-
luation statistique après lésion, et qui ont
des lésions profondes en C 3, montrent
des changements isignificatifis dans la
structure de la toile quand on compare
les toiles d’un même animal avant et
après lésion. Aucune des Araignées su-
bissant le même protocole opératoire,
mais ne recevant pas le rayonnement la-
ser, ne présente de changement dans la
toile. On en conclut que les changements
comportementaux sont liés à l’exposition
au laser.
La position de la lésion joue un rôle
essentiel dans la modification du compor-
23
Figure 4
Coupe sagittale de cerveau non lésé
Région postérieure (C 3 a) — Cm corps central.
Figure 5
Coupe sagittale de cerveau non lésé
Région antérieure (C 3 d)
Figure 6
Coupe sagittale de cerveau lésé
Région postérieure (C 3 a)
Les flèches indiquent Figure 7
remplacement Coupe sagittale de cerveau lésé
des cellules lésées Région antérieure (C 3 d)
tement. ‘Chaque fois que la lésion est
profonde et en C 3, un changement de
structure est observé dans la toile. Ceci
contraste avec le résultat obtenu avec 22
Araignées où les lésions sont en dehors de
la zone C 3. Parmi celles-ci, seules 14
Araignées construisent des toiles pertur-
bées. La probabilité, d’une telle modifi-
cation de structure après lésion en C 3
est significativement plus élevée que pour
des animaux avec des lésions situées en
une autre région (x2J P < .0)1). De plus,
il y a une plus grande cohésion dans les
changements occasionnés par les liions en
G3 que dans l’autre groupe.
Comme nous n’avons pas déterminé
avec précision la position des lésions hors
de la Zone C 3, nous ne pouvons qu’en-
visager la cause possible des changements
occasionnés dans la toile. Par exemple,
l’estomac pourrait être lésé dans sa fonc-
tion ce qui entraînerait un déficit alimen-
taire et, en conséquence, des toiles plus
petite®(Witt 1963). D’autre part, les
muscles, ou les attaches des muscles,
pourraient être atteints et la fonction
d’une ou de plusieurs pattes altérées. Con-
naissant le rôle important des premières
paires de pattes en particulier (Reed et
coIl.J1965 ; Le Guelte, 1965), cette even-
tualité n’est pas à écarter. Actuellement,
nos données ne pous permettent pas une
analyse plus poussée de ces résultats.
De nombreuses Araignées ne construi-
sent aucune toile après lésion en C 3. Le
nombre relativement restreint d’observa-
tions sur la fréquence de construction
après laser ne permet pas l’évaluation de
cette donnée, mais semble indiquer une
diminution après lésion en C 3.
Le tableau ne donne les résultats que
pour les changements dans les paramètres
de la toile significative à P<.0il (test de
Student). Cependant, toutes les toiles
construites par des Araignées lésées en
C 3 présentent des différences par rap-
port aux toiles avant lésion, dont beau-
coup ne sont pas hautement significatives.
La toile après lésion est toujours plus
circulaire qu’avant l’opération (la forme
ovale est modifiée par diminution de
l’axe vertical), les rayons sont significa-
tivement moins nombreux et les spirales
sont plus irrégulièrement posées. Les toi-
les sont plus petites, construites avec une
Araignée Position de la lésion (voir Fi$ l) Nombre de cellu- les lésées conséquences comportementales—
D 3 237 C 3 d 65 nombreux changements signif1- catifs à P < .02 pas de changement à F ^.01
IJ3 4. 14 C 3?êT ‘ 95 diminution du nbre de rayons (27/22ffljau nombre de spires régularité angulaire K^2/2jJS’.régularité des spires 0,8/l,3.
‘ D 4 23 C 3 d 60 surface de la zone libre (31 Am surface d’une maille (76/48) mm?
D 4 32 C 3 d 50 nombre de rayons 34/18 Nombre de spires 37/19 régularité angulaire 3» 01 /8,30
D 5 18 G 3 a 120 pas de toile après lésion
D 5 23a |PWi3 a 150 rayon* 21 *8 :16,0 |lf8 l$\Q,65 régularité angulaire 5,37/11,82
D 5 26a IFlIp3 a spires E 28/22 surface captrice 376/279 cmé surface zone libre 39/29 cm2 longueur de fil 16/l2m ‘Diminution .du nombre de rayons
D 5 42 e 3 c 55 spires E 27/20 surface captricéMO/310 cm2’,
D 5 47 C 3 c 155 . nombre de toiles insuffisant pour évaluation statistique
D 5 50 C 3 a 80 quotient d’excentricité 0,91/0,73 régularité angulaire 3*43/4,67
D 5 61 C 3 a 160 angle médian 15*/l 3# régularité angulaire 4»79 / 3>88
D 5 86 C 3 e 250 nombre de spires 19/l2 surface de la zone libre 38/l8 cm2
Seules les différences significatives
à P p . 01 sont indiquées.
longueur de fil inférieure et ont une sur-
face cap trice plus restreinte.
Bien que les variations dans la gravité
des modifications de la toile soient im-
portantes d’un animal à l’autre, un chan-
gement observé dans la première toile
après lésion chez un animal donné reste
constant tout au long du reste de la pé-
riode d’observation pour cet animal.
Discussion
Dans ces expériences, nous obtenons
des dégâts très comparables pour difïé-
25
rentes positions de la lésion dans le SNC
(voir tableau). Ceci s’oppose aux résul-
tats obtenus grâce à l’action de ‘drogues.
On sait, en effet, que chaque drogue pro-
duit un effet spécifique.
Les lésions affectant la zone du Corps
central (C 3 a) ne sont pas seules à mo-
difier le comportement de construction.
On sait (Meier, 1967) que cette région
est, au point de vue structural, une zone
de liaison entre fibres sensitives en pro-
venance des pattes locomotrices et fibres
motrices pour les mêmes appendices. Les
cellules plus antérieures (C 3 b, c, d,) ne
jouent pas de rôle dans la coordination
des mouvements des pattɧ|^La lésion des
fibres cheminant en dessous de ces cel-
lules, fibres provenant du Corps Central,
pourrait aussi avoir une importance dans
la création des désordres de la toile. Sans
envisager une localisation fonctionnelle
des cellules nerveuses, on peut toutefois
constater que les lésions au niveau C 3, c
perturbent la pose des spirales captrices
(déplacement circulaire de 1’(Araignée en
fin de toile) alors que les lésions en C 3, a
(corps central) intéressent en plus la pose
des rayons (déplacement||r linéaires de
l’Araignée au début de la construction).
Le nombre des cellules visiblement dé-
truites se situe entre 0,9 et 4 % du nom-
bre total des cellules du ganglion sus-
oesophagien. D’après les résultats que
nous obtenons sur les 12 Araignées avec
lésion en C3, une relation entre le nom-
bre de cellules lésées et la gravité des
désordres n’est pas mise en évidence. La
position de la lésion au niveau du corps
central |ui-même revêtirait une grande
importance. Il serait alors possible d’en-
visager, pour cette zone du SNC, un rô-
le prépondérant de centre coordinateur
sensori-moteur qui régulerait des élé-
ments de comportement moteur grâce aux
informations sensitives y arrivant. Ce qui
serait conforme à sa structure histologi-
que déjà connue.
Acknowledgements
During the long years of experimentation we
had the efficient help of Dr. Frank K. Tittel from
the Department of Electrical Engineering, Rice
University, Houston, Texas, and Dr. Charles F.
Reed, Department of Psychology, Temple Uni-
versity, Philadelphia, Pennsylvania. The work
would no|| have been carried out without the
valuable support from a grant of the National
Science Foudation and the support from the
North Carolina Department of Mental Health,
Division of Research.
Bibliographie
Babu (S.K.), 1965. — Anatomy of the central ner-
vous system of Arachnids. Zool. Jb. Anat. 82,
1-154.
Balesdent (M.L.), 068. 93|Microchirurgie par
rayonnemifflt laser : travaux sur ill
crustacé isopode As alius gggfeicus L. Bull.
Acad. Soc. Lorr. Scf. 7, 1, 82-92.
Legendre (R.), 1958. Contributions à l’étude
du système nerveux des Aranéides. Ann. Biol.
34, 5-6, 194-223.
Le Guelte (L.), 1965. — Répercussions de la perte
de pattes sur la construction de la toile chez
Araneus diadematus et Zygiella x-notata (Arai-
gnées, Argiopidae). Psychol. Fr. 10, 3, 257-
264.
Le Guelte (L.) et Witt (P.N.), 1968. — Données
morphologiques permettant de localiser sur
l’animal vivant les différentes régions du sys-
tèmes nerveux central de l’Araignée argiopide
Araneus diadematus Cl. soumise à des lésions
par action du laser. Bull. Mus. Nat. Hist. Nat.
40, 4, 742-744.
Meier (F.), 1967. — Beitràge zur Kenntnis der
postembryonalen Entwicklung der Spinnen Ara-
neida Labidognatha unter besonderer Berück-
sichtigung der Histogenèse des Zentral-ner-
vensystems. Rev. Suisse Zool. 74, 1, 127 p.
ReIm^FI Witt (P.N.)fc: JONES (R.L.) 1965 -U
The measuring function of the first legs of
Araneus diadematus Cl. Behaviour, 25, 12,
98-119.
Witt (P.N.) 1963. — Environnement in relation to
behavior of Spiders. Arch. Environ. Health 7,
4-12.
Wit|||(P.N.), Reed (C.F.) Peakall (D.B.), 1968. —
A spider’s web, problems in regulatory bio-
logy. Springer Verlag. Heidelberg. 107 p.
Witt (P.N.) 1969. — Behavioral consequences of
Laser lesions in the central nervous system of
Araneus diadematus Cl. Am. Zool. 9, 121-131.