Images Collection


Read OCR Digitized Article Text
NOTE: This plain text article interpretation has been digitally created by OCR software to estimate the article text, to help both users and search engines find relevant article content. To read the actual article text, view or download the PDF above.
Zeitschrift
tu’ vwtf
leichende Physiologie, ßd. 32, S. 29—45 (1950).
Aus dem Zoologischen Institut und dem Pharmakologischen Institut
der Universität Tübingen.
DIE BEEINFLUSSUNG DES NETZBAUES DER SPINNEN DURCH NEUROTROPE SUBSTANZEN.
Von
Hans M. Peters, Peter N. Witt und Dieter Wolfe.
Mit 6 Textabbildungen.
(Eingegangen am 13. Mai 1949.)
Eine Reihe neuerer Untersuchungen geben uns einen Einblick in die Gesetzmäßigkeiten, welche die Struktur und die Gliederung des Radnetzes der Spinnen beherrschen. Auch wissen wir in großen Zügen, wie die Spinne bei der Herstellung ihres kunstvollen Gebildes verfährt. Damit sind gewisse Voraussetzungen gegeben, die es gestatten, die Untersuchungen auf ein sehr schwieriges Problem auszudehnen, das bisher gänzlich außer Reichweite lag, nämlich auf die zentralnervösen Prozesse, welche dem Verhalten der Spinne zugrunde hegen. In früheren Arbeiten hatte sich das Spinnennetz als ein Modell erwiesen, an dem sich gewisse fundamentale Lebenserscheinungen, nämlich Gestaltungsprozesse, besser studieren lassen als an vielen anderen Objekten, weil in ihm eine Anzahl äußerst günstiger Untersuchungsbedingungen Zusammentreffen. Vielleicht gilt dies ebenso mit Bezug auf das neu auftauchende Problem der zentralnervösen Grundlagen von Instinktbewegungen.
Wir müssen uns wohl vorstellen, daß die Tätigkeit der Spinne beim Netzbau auf dem Zusammenwirken einer Anzahl verschiedener zentraler Grundfunktionen beruht. Wenn wir einen Einblick in dieses Zusammenspiel gewinnen wollen, müssen wir einen Weg finden, jene Funktionen voneinander zu trennen. Mechanische Eingriffe in das Zentralnervensystem erscheinen aus technischen Gründen äußerst schwierig, wenn nicht unmöglich. Aber vielleicht dürfte von einer Beeinflussung mittels neurotroper Substanzen ein Erfolg erwartet werden (Peters). Wenn wir einen Versuch in dieser Richtung unternommen haben, so geschah dies auch deshalb, weil wir uns im Spinnennetz einen guten Test zur Prüfung von Substanzen überhaupt versprachen (Witt). Es ist nämlich zu erwarten, daß ein Gebilde, das durch Integration so mannigfaltiger Funktionen entsteht, ein sehr feines Reagens auf Substanzen darstellt. Dieses Reagens bietet überdies den großen Vorteil, daß es die Substanzwirkung automatisch aufschreibt und in einem Dokument festhält, das sehr genau analysiert werden kann.
Hans
M. Peters, Peter N. Witt und
Dieter Wolfe:
Wir hoffen mit dem weiteren Ausbau der Methodik auch in ihrer Wirkung unbekannte Medikamente, soweit sie das Zentralnervensystem beeinflussen, durch ihre Auswirkung auf den Netzbau der Spinne charakterisieren zu können. Jede Analyse der am Netzbau beteiligten Funktionen kommt der pharmakologischen Auswertung des Testes zugute, und jede Beobachtung der Wirkung bekannter Substanzen auf das Netz dient der Vermehrung unserer Kenntnisse über die am Netzbau beteiligten Funktionen, so daß sich in unserer Arbeit zoologische und pharmakologische Interessen ergänzen.
Unsere ersten Ergebnisse lassen die begonnenen Untersuchungen aussichtsreich erscheinen. Die Methode ist nach den verschiedensten Seiten ausbaufähig, doch ist das Feld, das sich der experimentellen Arbeit darbietet, so groß, daß es unsere eigenen Arbeitsmöglichkeiten übersteigt. Wenn wir heute schon Beobachtungen mitteilen, so geschieht dies hauptsächlich in der Erwägung, unsere Methode könnte vielleicht anderen von Nutzen sein. Mit Rücksicht darauf möchten wir unsere ersten Befunde schon jetzt veröffentlichen, obwohl dies zunächst nur in einer vorläufigen Form geschehen kann.
Nach den ersten Vorarbeiten erwies es sich als erforderlich, eine zuverlässige quantitative Methode der Einverleibung von Substanzen auszuarbeiten. Diese Aufgabe hat Wolff übernommen L
Material und Technik1 2.
Als Versuchstier diente die Radnetzspinne Zilla x-notata (= litter ata). Sie gehört zur Familie der Araneidae, also in die Verwandtschaft der Kreuzspinnen. Wir wählten diese Art, weil sie sich leicht im Laboratorium halten läßt und sehr rege im Netzbauen ist. Auch ist das Netz dieser Spinne so genau bekannt, daß eine genügend breite Basis für die Beurteilung von Substanzwirkungen gegeben war. Eingehende Untersuchungen von Peters über die Feinstruktur des Zilla-Netzes stehen vor ihrem Abschluß.
Die jüngeren Entwicklungsstadien von Zilla sind wegen ihrer Kleinheit für pharmakologische Experimente ungeeignet. Wir benutzten nur Tiere nach der vorletzten und letzten Häutung.
Haltung der Versuchstiere.
Zilla x-notata ist eine unserer häufigsten Spinnen. Man findet sie an Gartenzäunen, Kellerfenstern, in Ställen u. dgl. in großen Mengen, so daß Beobachtungsmaterial jederzeit zu haben ist. Die jungen Spinnen schlüpfen im Frühjahr aus überwinterten Eiern und werden nach einigen Häutungen im Herbst geschlechtsreif. Danach sterben sie. Tagsüber halten sie sich in einem Schlupfwinkel auf, wobei sie mit der Warte des Netzes durch einen Signalfaden verbunden sind (Abb. 1). Diese Lebensweise macht die Art für Versuche besonders geeignet. Bietet man den Spinnen in einem Rahmen aus Holzleisten einen passenden Schlupfwinkel, so kann man sie leicht darin ansiedeln und lange Zeit dort halten.
1 Für die Mithilfe bei der Auswertung sind wir Dr. Erika Bierbrodt zu großem Dank verpflichtet.
2 Eine allgemeine Orientierung über die Biologie der Spinnen geben Gerhardt und Kästner (1938). Über Radnetze vgl. man die Arbeiten von Wiehle, Tilquin, Peters.
4
t
30
Die Beeinflussung des Notzbaues der Spinnen,
31
Als Schlupfwinkel fertigten wir kleine konisch geformte Papierdüten an, die wir in einer der oberen Koken des Kähmens befestigten. Je nach dem Alter der Spinnen wählt man quadratische Kähmen von 25—50 cm Kantenlänge.
Die Übertragung einer frei lebenden Spinne in einen Rahmen geschieht folgendermaßen: Man lockt das Tier mit einer Beute in das Netz. Während es sieh dort befindet, befestigt man die Düte so an das obere Ende des Signalfadens,

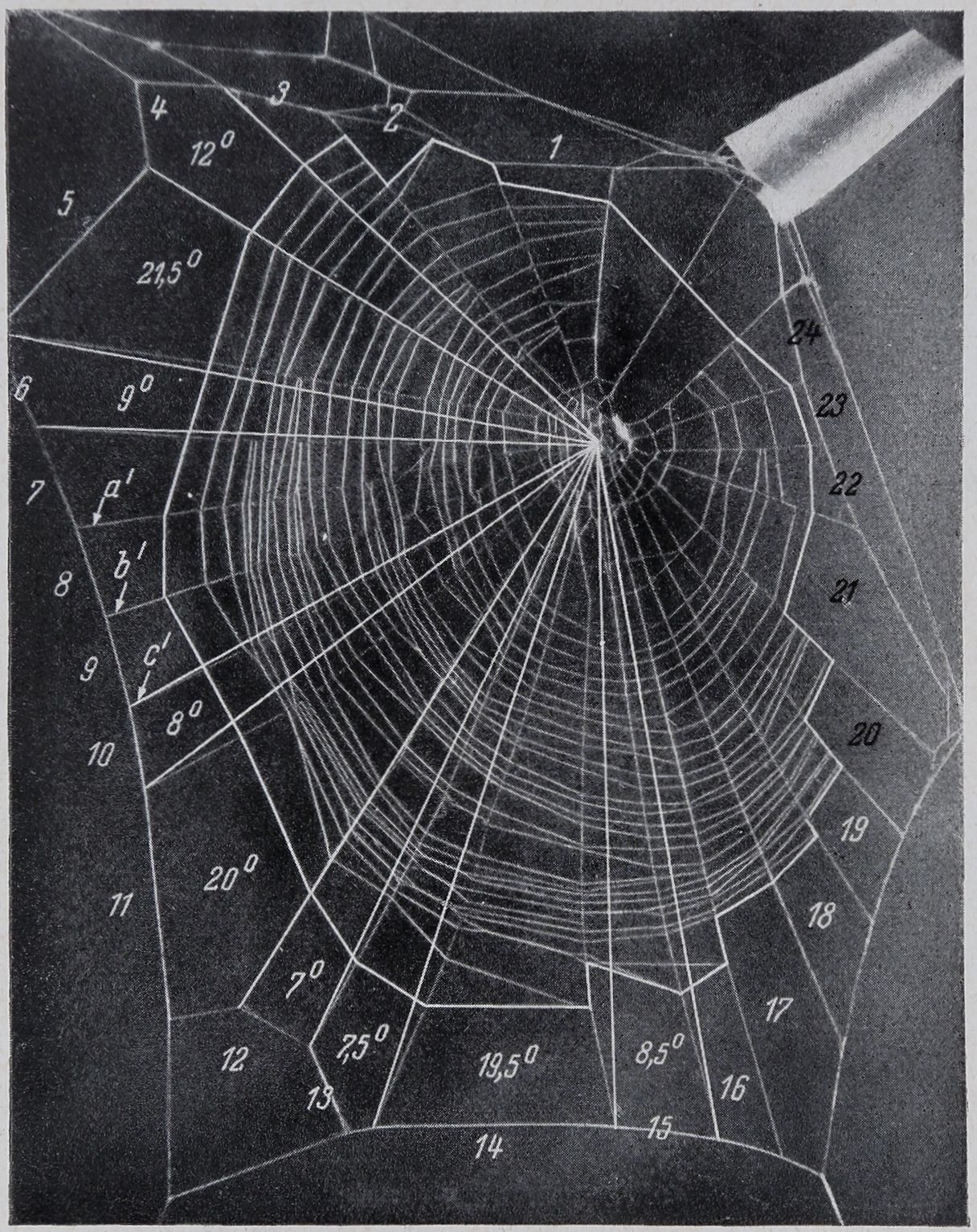
Abb. 1. Netz einer Geschlechtsreifen Zilla x-notata. 1 Nabe; 2 Befestigungsumgänge, beides zusammen die „Warte“; 3 freie Zone; 4 freier Sektor; 5 Signalfaden; 6 Schlupfwinkel; 7 Fangbereich (mit Klebfäden); 8 Radialfäden; 9 Rahmenfäden; 10 Spannseil. Die graphische Darstellung Abb. 5 a bezieht sich auf die Abstände der Klebfäden längs
der Radialfäden a, b und c. 1: 2,5 nat. Größe.
daß die Spinne bei ihrer Rückkehr in die neue Wohnung hineinlaufen muß. Um sie an den Schlupfwinkel zu gewöhnen, beläßt man diesen einen Tag an Ort und Stelle. Dann überträgt man ihn mit der Spinne auf den Rahmen. Im allgemeinen baut das Tier sein Netz vom neuen Platz aus. Die Rahmen hängt man zweckmäßig an ein Gestell aus Drähten an der Fensterseite eines Zimmers auf. Man kann so auf kleinem Raum eine große Anzahl von Spinnen unter Beobachtung halten. Hin und wieder kommt, es vor, daß Spinnen abwandern, aber im allgemeinen sind sie sehr seßhaft. Es muß auch damit gerechnet werden, daß eine umherwandernde Spinne eine andere aus ihrem Schlupfwinkel vertreibt; unter Umständen ist also eine individuelle Markierung der Versuchstiere geboten.

Hans M. Peters,
Peter N. Witt und
Dieter Wolfe :
Fattening. Wenn eine Heute in das Netz gerät, so eilt die Spinne am Signalfaden herbei, läuft über die Warte zur Fangstelle, tötet das Opfer durch einen Biß, spinnt es ein und trägt es in den Schlupfwinkel. Erst da verzehrt sie ihren Fang. Daß die Spinne sich tagsüber in ihrem Schlupfwinkel aufhält, wurde schon gesagt. Nur während der Dunkelheit befindet sie sich in der Warte des Netzes. Gefangene Spinnen kann man mit Drosophila füttern. Für größere Tiere genügen
zwei Fliegen täglich. Trockenheit der Luft soll vermieden werden. Unter Umständen gibt man ein Stückchen Fließpapier mit Wasser an den Schlupfwinkel, so daß die Spinne nach Belieben Wasser aufnehmen kann.
Netzhau. In der Regel erneuert Zilla ihr Netz täglich, wobei sie Teile des alten Netzrahmens verwendet. Um auf jeden Fall ein neues Netz zu erhalten, zerstört man am Abend das alte bis auf wenige Fäden. Man schneidet einfach einige der Spannfäden (Abb. 1), mit denen das Netz am Holzrahmen befestigt ist, mit der Schere durch, so daß sich das Gewebe zusammenzieht. Die Spinne frißt vor der Herstellung eines neuen Netzes die Reste des alten auf. Wenn man das alte Netz vollständig entfernt, wandert die Spinne nicht selten ab. Obwohl Zilla gelegentlich auch am Tage baut, ist die Netzbauzeit doch recht genau auf die frühen Morgenstunden, noch vor der Morgendämmerung, festgelegt. Uber die Abhängigkeit der Netzbauzeit von verschiedenen Außeneinflüssen liegen Untersuchungen von Spbonk (1935) an Zilla und an der Kreuzspinne vor. Hat eine Spinne mit dem Netzbau begonnen, so läßt sie sich durch künstliche Beleuchtung nicht stören. Man kann sie daher gut bei ihrer Tätigkeit beobachten.
Häutung. In den Zeiten der Häutung setzt die Spinne mit dem Netzbau aus. Schon einige Tage vor der Häutung selbst sind die Netze nicht selten unregelmäßig und verkleinert. Bei der Auswertung von Versuchsergebnissen ist dies naturgemäß zu berücksichtigen.
Nähere Angaben finden sich bei Tilqtjin (1938). Die Exuvien gestatten eine genaue Messung der Größe der Versuchstiere in der vorausgegangenen Phase, was unter Umständen von Bedeutung ist. Die Haut wird in Kalilauge erhitzt, mit Wasser ausgewaschen, über die Alkoholreihe entwässert und über Xylol in Canadabalsam eingeschlossen.
Das Photographieren der Netze. Genaue Netzmessungen, die zur Analyse der Effekte unumgänglich sind, lassen sich nur an Photographien ausführen. Zum Photographieren bedienten wir uns eines von H. Homann angegebenen Verfahrens K Es besteht darin, daß man die Netzfäden mit Ammoniumchlorid anräuchert, wodurch sie auf der Photographie deutlich sichtbar werden.
In einem dicht schließenden Kasten mit einem abnehmbaren Deckel werden Schalen mit rauchender Salzsäure und verdünntem Ammoniak aufgestellt. Die Salzsäure befindet sich jeweils in einer größeren Schale, in welche die Schale mit Ammoniak hineingestellt wird. Nach etwa einer Viertelstunde hat sich in der Regel der nötige Belag niedergeschlagen. Obwohl die Spinnen gegen die Dämpfe verhältnismäßig unempfindlich sind, empfiehlt es sich, sie ihnen nicht länger als notwendig auszusetzen.
Bei der photographischen Aufnahme ist darauf zu achten, daß die Proportionen des Netzes richtig wiedergegeben werden. Ein an den Holzrahmen befestigter Maßstab wird mitphotographiert, so daß die Negative auf ein bestimmtes Größenverhältnis kopiert werden können. Das Netz wird schräg von hinten, von beiden Seiten, mit je einem 500-Watt-Scheinwerfer beleuchtet. Eine schwarze Wand bildet den unbeleuchteten Hintergrund.
1 Für die briefliche Mitteilung seiner außerordentlich brauchbaren Methode sind wir Herrn Dr. Homann zu großem Dank verpflichtet.
Dio Boo inf lu bB Jig des Notzbaues der Spinnen,
Applikation und Dosierungl. Es wurde versucht, die Applikation möglichst sauber, einfach und natürlich zu gestalten, d. h. die Bedingungen zu finden, bei denen die Substanz unter Erhaltung eines physiologischen Ablaufes der Nahrungsaufnahme möglichst rein angeboten werden konnte.
Grundlegend ist die Beobachtung, daß die Spinnen Wasser jederzeit begierig auf sau gen.
Attrappen, wie leere Fliegenpuppen, Glaskugeln und Kügelchen aus Holundermark, Wattestückchen, Paraffinbehälter und Röhrchen aus Bienenwachs mit Fliegenteilen wurden von der Spinne entweder sofort oder nach einiger Zeit aus dem Netz entfernt, oder nach kurzem Betasten und Bearbeiten mit den Mundteilen hängen gelassen. Nur selten beschäftigten die Tiere sich längere Zeit (2—3 Minuten) mit der Attrappe.
Wenn diese allerdings Wasser oder Bouillon enthielt, begannen die Tiere zu saugen. Versuche mit Fliegenbouillon ergaben im allgemeinen eine zögernde Annahme über einige Minuten, niemals jedoch volle Annahme mit Abtransport in den Schlupfwinkel. Bei einem Versuch mit gezuckerter Bouillon in einem ausgehöhlten Fliegenabdomen wurden überraschenderweise Einspinnen und Abtransport beobachtet, was sich dann in weiteren Versuchen mit Zuckerwasser (Rohrzucker) regelmäßig wiederholte.
Entsprechende Beobachtungen anderer Autoren über positive Zuckerwirkung bei Spinnen sind uns nicht bekannt. In weiteren Versuchen erwies sich der Zucker zugleich als ein brauchbares Geschmackscorrigens.
Aus den geschilderten Vorversuchen ergab sich folgende Methode der Applikation :
Aus einer Zucht der Schmeißfliege (Lucilia caesar) oder einer ähnlichen Art wird eine Anzahl Tiere mit Äther getötet. Die Hinterleiber werden abgeschnitten und auf einer Glasplatte unter Verwendung von Pinzetten und einer feinen Nadel ausgequetscht. Es bleibt nur das ausgehöhlte Chitinskelet zurück, das einige Stunden an der Luft getrocknet wird. Die Attrappen werden mit der gezuckerten Substanzlösung gefüllt. Bei bitteren Substanzen erweist es sich am günstigsten, die Lösung so stark zu zuckern, daß sie sehr süß schmeckt. Nachdem die Attrappe gewogen ist (sie wiegt 15—30 mg), wird sie ins Netz gegeben. Dann lockt man die Spinne mit einer Stimmgabel (etwa 835 Hertz) in das Netz, indem man die Warte einen Augenblick berührt. An Stelle einer Stimmgabel kann man auch eine lebendige Fliege verwenden, die man kurze Zeit an das Netz hält und beim Erscheinen der Spinne wieder fortnimmt. Die Spinne saugt eine gewisse Zeit an der Attrappe, die man in den meisten Fällen selbst bestimmt; denn nur selten — bei bestimmten Substanzen und vielleicht bei zu starken und zu schwachen Zuckerkonzentrationen— hört die Spinne selbständig nach etwa 5—10 Minuten zu saugen auf. In 30 Minuten haben die Tiere im allgemeinen mehr als die Hälfte getrunken. Dann werden die Attrappen mit einer Pinzette der Spinne wieder fortgenommen und abermals gewogen. Aus der getrunkenen Menge (Gewichtsverlust), der Konzentration der verwendeten Substanzlösung und derem spezifischen Gewicht (Zucker) läßt sich die aufgenommene Menge der Testsubstanz berechnen. Erfahrungsgemäß trinkt eine mittelgroße Zilla in einer halben Stunde zwischen 2—10 mg Lösung. Indem man die Konzentration der Substanz im Lösungsmittel variiert, läßt sich die zu verabfolgende Menge vorher ungefähr dosieren.
Die verschiedenen Phasen der Substanzwirkung lassen sich feststellen, indem man den zeitlichen Abstand zwischen Applikation und voraussichtlichem Beginn des Netzbaues verschieden wählt. Wie erwähnt, baut die Spinne in den frühen
1 Vgl. zu Folgendem: Baltzer (1923), Millot (1946).
Zeitschr. f. vergl. Physiol. Bd. 32.
3
34
Hans M. Peters, Peter N. Witt und Dieter Wolff:
Morgenstunden, und zwar schon vor Sonnenaufgang. Nach der Jahreszeit liegen die Bauzeiten verschieden.
Die Methode der Dosierung wurde erst mit dem Fortgang der Untersuchungen ausgearbeitet. In den gleich zu schildernden Experimenten haben wir noch nicht, oder nur ungenau, dosiert. Die Substanzlösung wurde den Spinnen an einem mit ihr getränkten Stückchen Fließpapier geboten oder ihnen einfach als Tropfen an die Mundteile gehängt.
Auswertung der Versuchsergebnisse. Wie schon erwähnt, wurden die Netze zur Auswertung der Versuche photographiert. Wir machten es uns zur Regel, grundsätzlich jedes Netz einer unter Beobachtung gehaltenen Spinne zu photographieren, auch wenn es nicht unter Substanz Wirkung hergestellt worden war. Auf diese Weise hatten wir zur Beurteilung der Substanzeinflüsse eine ausreichende Kontroll-möglichkeit an normalen Netzen. Am klarsten läßt sich ein Substanzeffekt nachweisen, wenn nach einer Reihe normaler Netze unbeeinflußter Spinnen Netze auftreten, welche nach einer Gabe von Substanz den charakteristischen Einfluß derselben zeigen. Im folgenden geben wir eine Auswahl von Ergebnissen der laufenden Untersuchungen. An ihnen wollten wir lediglich die Brauchbarkeit und Ausbaufähigkeit der Methode zeigen. Eine genauere Analyse der Effekte behalten wir späteren Veröffentlichungen vor.
Wir unterscheiden die Substanzwirkungen nach Art und Schwere des Effektes. Zunächst untersuchen wir die Frage, ob es möglich ist, den Bautrieb als solchen zu hemmen oder seine Auslösung zu fördern. An zweiter Stelle soll geprüft werden, ob es möglich ist, die Größe des Netzes zu beeinflussen und schließlich untersuchen wir die Beeinflußbarkeit der Netzgestalt selber, wobei wir zwischen Grundstruktur und Feinstruktur unterscheiden (Peters 1937, 1939). 1 2
1. Hemmung des Bautriebes.
Beeinflussung mit Veronal. Mit Veronal versuchten wir den Netzbau zu hemmen. Die Substanz wurde 6mal in einer Konzentration 1:100 und 9mal in einer Konzentration 1:1000 6—8 Stunden vor der Netzbauzeit gegeben, insgesamt also lömal an verschiedenen Individuen. Die Bauzeit als solche wurde von der Substanz anscheinend nicht beeinflußt. Die Dosis lag in der Größenordnung von etwa 5—50 y.
1. Konzentration 1:100. Effekt: 4mal Ausfall des Netzbaues für 1 Tag; lmal deutlich verkleinertes Netz am nächsten Tag; lmal ein ganz abnormales, sehr kleines Netz am nächsten Tag.
2. Konzentration 1:1000. Effekt: 3mal Ausfall des Netzbaues für 1 Tag; 6mal verkleinertes Netz am nächsten Tag.
Zur Beurteilung der Effekte ist es wichtig zu beachten, daß die 7 Tiere, welche nach Veronal mit dem Netzbau aussetzten, mindestens noch am Tage vor dem Versuch ein Netz gebaut hatten. Einige von
Dio Beeinflussung dos Notzbuuos dor Spinnen. Bo
ihnen standen sohon länger unter Kontrolle. Eine hatte 14 Tage lang ihr Netz täglich erneuert. Die durchschnittliche Anzahl aufeinanderfolgender Tage, an denen jene 7 Spinnen festgestelltermaßen gebaut hatten, beträgt 5,3.
* o
Nach diesem Ausfall der Versuche erscheint es uns sehr wahrscheinlich, daß Veronal den Bautrieb hemmt.
Abb. 2. Netz der gleichen Spinne wie Abb. 1. 11 Tage später nach einer Gabe von etwa 6 y Pervitin etwa 6 Stunden vor Beginn des Netzbaues. — Die stark ausgezogene Linie gibt die Ausdehnung des Fangbereiches an. 1: 2,5 nat. Größe.
Weitere Erklärung im Text.
Zur Prüfung der Frage, ob auch eine Förderung des Bautriebes durch Substanzen möglich ist, eignen sich Spinnen in der letzten Lebensphase im Herbst. Die Intensität ihres Bautriebes läßt gegen Ende ihres Lebens immer mehr nach. Wir haben Fälle, die uns vermuten lassen, daß man in dieser Lebenszeit mittels Strychnin den entschwindenden Bautrieb wieder anregen kann (Ursula Hempel).
Beeinflussung der Netzgröße. Obwohl unsere Untersuchungen in dieser Frage noch in den Anfängen stecken, läßt sich doch schon mit Wahrscheinlichkeit sagen, daß man die Netzgröße durch eine Reihe von Substanzen unter das Normalmaß herabdrücken kann.
Zeitschr. f. vergl. Physiol. Bd. 32. 3a

Hans M. Peters, Peter N. Witt und Dieter Wolfe:
36
Als „Größe“ des Netzes bezeichnen wir die Ausdehnung der Netzfläche innerhalb der äußersten Umgänge der Klebfäden (Abb. 2, eingezeichnete Grenzlinie). Zur Messung wurde die bezeichnete Umrißlinie von den Photographien (die alle in dem gleichen Maßstab im Verhältnis zur natürlichen Größe des Netzes kopiert worden waren) auf Papier immer der gleichen Sorte durchgezeichnet. Die Figuren
wurden ausgeschnitten und gewogen. Dann konnte von dem Gewicht auf das Flächenmaß umgerechnet werden. Es sei noch erwähnt, daß im freien Sektor die Umrißlinie durch eine eingezeichnete Linie ergänzt wurde (vgl. Abb. 2).
Wir wählen als Beispiel das Pervitin. Die Säulen in den beiden Diagrammen (Abb. 3) stellen die Größe von Netzen dar, die an aufeinanderfolgenden Tagen gebaut wurden. Pfeile bedeuten die Verabfolgung von Pervitin1 6—7 Stunden vor Beginn des Netzbaus. Mit einer Ausnahme führte die Pervitingabe jedesmal zur Verkleinerung des Netzes. Die Pervitin-netze sind die beiden kleinsten in der Serie der Netze von Spinne Nr. 4 (obere Reihe). Auch das kleinste Netz von Nr. 1 (untere Reihe) ist ein Pervitinnetz. Allerdings ist unter den dreien in der Größe folgenden Netzen dieser Serie ein anderes kleines Netz, das nicht unter Substanz -ein Wirkung her gestellt worden ist. Die mittlere Größe der 6 Pervitinnetze beträgt 23 di 2 Einheiten2, die der 17 Kontrollnetze 30 ± 1,1 Einheiten. Es handelt sich sicher nicht um substanzspezifische Wirkungen, da pharmakologisch so verschiedene Mittel wie Veronal, Morphium, Cardiazol, Pervitin anscheinend vergleichbare Effekte hervorrufen. Die Diagramme zeigen, daß die Netzgröße schon normalerweise und ohne erkennbare Ursachen in weiten Grenzen schwankt. Das macht eine sichere Beurteilung der Substanz Wirkung schwierig. Vor der Inangriffnahme weiterer Experimente wird es nötig sein, die natürliche Variation der Netzgröße bei Zilla genau zu untersuchen.
Beeinflussung der Grundstruktur. Mit Coffein gelang es, die Netzgestalt als Ganzes weitgehend zum Zerfall zu bringen. Abb. 4 zeigt
Abb. 3. Die Netzgrößen zweier Zilla litterata an aufeinanderfolgenden Tagen. Die Pfeile bedeuten Pervitin-Gaben vor Beginn des Netzbaues.
1 Pervitinlösung I : 1500 etwa 4 3mg (etwa 6 y Substanz).
2 1 Einheit = 3,75 cm2, 23 Einheiten = 86,25 cm2, 30 Einheiten
11
;> ein


Dio Beeinflussung dos Netzbaues dor Spinnen,
37
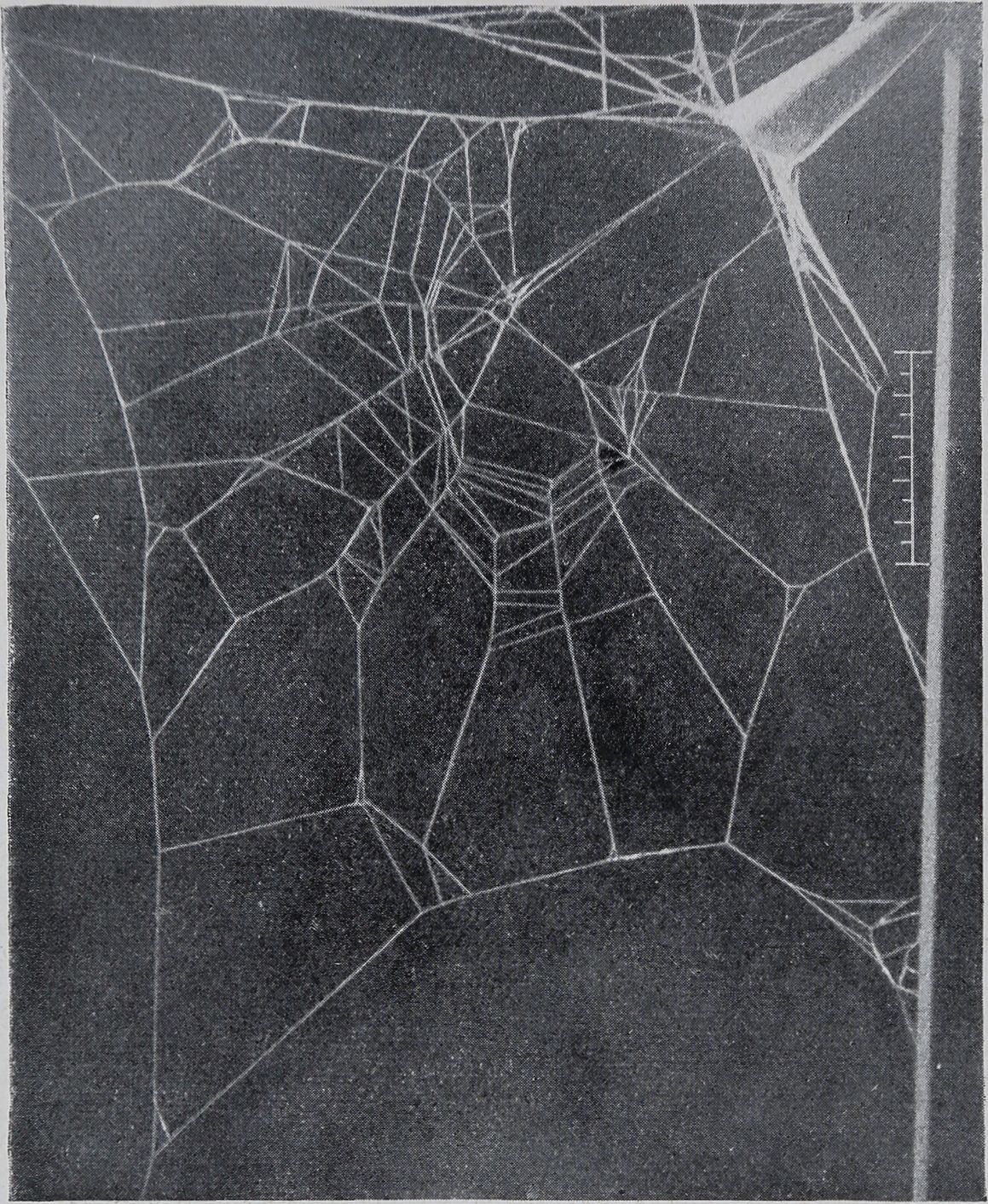
das Netz einer Spinne nach Applikation von etwa 50 y Coffein etwa 5 Stunden vor Beginn des Netzbaues. Dieses Netz weist alle Züge auf, die für den Typus des Zilla-Netzes charakteristisch sind, aber in einer teilweise sehr starken Unregelmäßigkeit der Anordnung. Die exzentrische Verlagerung des Mittelpunktes in Richtung auf den Schlupfwinkel entspricht dem normalen Netz. Ebenso ist der für Zilla charakteristische

Abb. 4. Netz einer Zilla x-notata nacb Coffeingabe (etwa 50 y etwa 5 Stunden vor Beginn
des Netzbaues). 1 : 3,3 nat. Größe.
„freie Sektor“ vorhanden. Der Netzrahmen scheint ebenfalls normal. Die Anzahl der Radialfäden ist auffallend gering, die Warte nur durch einige wenige konzentrische Fäden angedeutet. Die Klebfäden sind in einer außerordentlich unregelmäßigen Weise angeordnet. Das ganze Gebilde wirkt wie ein grobes Schema des Zilla-Netzes, in dem zwar alles Charakteristische enthalten ist , aber in einer besonders die feinere Ausgestaltung betreffenden außerordentlich gestörten Weise. Unterscheiden wir mit den Gestaltspsychologen (Sander) in der Aktualgenese der Gestalten Vorgestalt und Endgestalt, so ist das in Rede stehende Netz auf dem Stadium der Vorgestalt stehengeblieben. Die Spinne benötigte zur Herstellung dieses Netzes eine anomal lange Zeit, nämlich 23/4 Stunden, gegenüber der Normalzeit von etwa 1 Stunde.
Hans M. Peters, Peter N. Witt und
Dieter Wolef:
Mehr oder weniger große Störungen beobachteten wir in einer Anzahl weiterer Versuche mit Coffein. Das angeführte Beispiel stellt den bei weitem äußersten Fall dar1.
Beeinflussung der Feinstruktur. Die Feinstruktur des Netzes, wie sie besonders in der Maschung des Fangbereiches zum Ausdruck kommt, ist ein sehr empfindliches Reagens auf die verschiedensten Substanzen. Auch wenn die Netze sonst kaum beeinflußt erscheinen, kommt es doch sehr leicht zu einer Störung in der Anordnung der Klebfäden. Solche Beeinflussung beobachteten wir nach Applikation z. B. von Pervitin und Morphium hydrochloricum.
Wir beschränken uns in dieser Mitteilung auf eine Besprechung der Pervitin Wirkung. Eine feinere vergleichende Analyse wird wahrscheinlich eine mehr oder weniger große Spezifität der Substanzen erweisen2.
Beeinflussung durch Pervitin. Das folgende gründet sich auf 36 Versuche an 10 verschiedenen Individuen von Zilla x-notata und die Untersuchung einer großen Anzahl von normalen Kontrollnetzen. Abb. 1 zeigt das normale Netz einer Zilla x-notata und Abb. 2 ein Netz des gleichen Tieres 11 Tage später nach einer Gabe von etwa 6y Pervitin etwa 6 Stunden vor Beginn des Netzbaues. Gegenüber dem Normalnetz fällt vor allen Dingen eine erhebliche Unregelmäßigkeit in der Anordnung der konzentrischen Umgänge der Klebfäden auf. Gestört ist 1. die Regelmäßigkeit des Kurvenverlaufs der einzelnen Fadenzüge, 2. die Regelmäßigkeit der Abstände von Umgang zu Umgang. Im Normalnetz folgen die Abstände von Klebfaden zu Klebfaden von der Peripherie nach dem Zentrum hin einer strengen Gesetzmäßigkeit. Für die Kreuzspinne Aranea diadema ist diese von Peters genauer formuliert worden3. Die diesbezüglichen Untersuchungen am Zilla-Netz, welche zu einem grundsätzlich gleichen Ergebnis geführt haben, stehen vor dem Abschluß.
Die Anordnung der Klebfäden in einem bestimmten Sektor des Netzes läßt sich graphisch anschaulich darstellen, indem man die längs der Radialfäden zentralwärts aufeinanderfolgenden Abstände in ein Koordinatennetz einträgt. In Abb. 5a ist dies zunächst für ein unbeeinflußtes Zilla-Netz geschehen. Wir wählten zu unseren Messungen jeweils 3 benachbarte Radialfäden, um zugleich einen Einblick in die natürliche Streuung der fraglichen Werte zu gewinnen. Der Vergleich einer solchen Darstellung mit einem entsprechenden Diagramm aus einem Pervitinnetz zeigt die außerordentliche Vergrößerung der Streuungsbreite bei dem letzteren (Abb. 5b). Dies ist einer unserer sichersten und regelmäßigsten Effekte. An Stelle der gegebenen Beispiele
1 Anmerkung bei der Korrektur: Inzwischen von Witt mehrmals reproduziert..
2 Den ersten Versuch einer Klärung unternahm Witt auf der Tagung des Schweizer Vereins der Physiologen und Pharmakologen (22./2S. 10.49): „Verschieden© Wirkung von Pervitin und Coffein auf den Netzbau der Spinnen“. Im Druck.
3 Peters: %. Naturforsch. 2b (1947).
Dio Beeinflussung dos Netzbaues der Spinnen,
39
könnten wir zahlreiche andere anführen, alle mit den charakteristischen Abweichungen gegenüber Normalnetzen, Ein genauer quantitativer Vergleich der normalen mittleren Streuung mit der Streuung nach Pervitin steht noch aus.
Wir möchten den Effekt vorläufig so interpretieren, daß ihm im Wesentlichen eine Störung der sensorischen Kontrolle zugrunde liegt. Normalerweise bestimmt die Spinne den Abstand von Klebfaden zu
Abb. 5 a u. b. a Streuung der Klebfadenabstände in einem normalen Zilla-Netz (Abb, 1, gemessen längs der Radialfäden a, b, c); b Streuung unter Pervitinwirkung. (Abb. 4, gemessen längs der Raaialfäden a’, b’, e’.)
Klebfaden durch Abtasten mit einem Vorderbein (Peters 1939). Sie beginnt bei der Herstellung der Eangfäden außen an der Peripherie und schreitet dann in Rundgängen oder Pendelzügen allmählich gegen das Zentrum vor. Nach Pervitin erscheint die Kontrolle der Abstände an den bereits hergestellten Umgängen weitgehend gestört. Die Tastbewegung ist zwar noch erhalten, aber es kommt oft vor, daß die Spinne sozusagen ins Leere greift oder auch trotz Berührung des Kontrollfadens einen anomal großen oder kleinen Abstand wählt. Dies geht auch aus einem Film hervor, in welchem die Bewegung einer Kreuzspinne nach Pervitingabe aufgenommen worden ist1.
Auch hinsichtlich der Anordnung der Radialfäden sind die Pervitin-netze unregelmäßiger als normale. Einiges darüber wird im folgenden Kapitel mitgeteilt.
1 Hergestellt in Verbindung mit dem Institut für Film und Bild (Göttingen).
Zeitschr. f. vergl. Physiol. Bel. 32. 3b


40
Hans M. Peters, Peter N. Witt und Dieter Wolff:
Ausfallerscheinungen. Ein besonders feiner Test betrifft gewisse Ausfallerscheinungen. Die Beobachtungen gründen sich auf die sogenannten Restsektoren (vgl. dazu Peteks 1937). Zum Verständnis des

Abb. 6. Netzgerüst einer halbwüchsigen Kreuzspinne. Die arabischen Zahlen bezeichnen die Reihenfolge der Radialfäden. Ein < hinter der Zahl bedeutet, daß die Speiche zusammen mit dem betreffenden Rahmenfaden hergestellt wurde. Die Reihenfolge der Rahmenfäden ist durch römische Zahlen bezeichnet. Faden 8 ist ein Radius, der wahrschéinlich später durch 13 ersetzt wurde. Der allererste Faden dieses Netzes ist mit a bezeichnet. Dann wurde der Rahmenfaden I hergestellt, dann Radialfaden 2 mit Rahmenfaden II usw. Die Pfeillinien am untersten Rahmen bedeuten Laufgängo der Spinne zur Verstärkung dieses Fadens. Für Zilla gilt grundsätzlich die gleiche Gesetzmäßigkeit in der Herstellung der Radialfädeh wie für die Kreuzspinne. Nach
einer Photographie.
Folgenden müssen wir darauf eingehen, wie die Spinne bei der Herstellung ihrer Radialfäden verfährt (Abb. 6). Zu Beginn des Netzbaues stellt sie zugleich mit einem Radialfaden einen (primären) Rahmen-faden her, also z. B. den Radialfaden 4 in einem Arbeitsgang mit dem
DU’ Beeinflussung dos Netzbaues der Spinnen.
41
langen Rahmeufadon auf der reohten Seite des Netzes. Später werden sogenannte sekundäre Rahmenfäden eingesetzt, ebenfalls zugleich mit je einem Radialfaden, also z. B. Radialfaden 19, Radialfaden 20, Radialfaden 17, Radialfaden 2 und Radialfaden 12 mit den betreffenden sekundären Rahmenfäden. Es entsteht auf diese Weise ein Netzgerüst aus einem Rahmenwerk und einer Anzahl nach den verschiedenen Richtungen ausstrahlender Speichenfäden. Bezüglich der Einfügung der weiteren Radialfäden herrscht nun die Regel, daß die Spinne sie in dem für den betreffenden Netzabschnitt geltenden Winkelabstand jeweils von oben nach unten an die bereits vorhandenen Radialfäden anfügt. Sie läuft also etwa vom Zentrum aus an dem Radialfaden 4 zur Peripherie, dann ein Stückchen abwärts und befestigt den neuen Faden bei 5, worauf sie an ihm in das Netzzentrum zurückkehrt. Als nächsten stellt sie den Radialfaden 6 her, wobei sie die Speiche 1 als Laufsteg benutzt. Bei der Herstellung des Fadens 7 benutzt sie den Radial -faden 2, dann zieht sie den Faden 8, läuft in entgegengesetzter Richtung am Faden 5 wieder zum Rahmen und stellt die Speiche 9 her. Auf den Faden 9 folgen später 11, 14 und 15. Auf 15 folgt nach einiger Zeit die Speiche 19, wobei diese aber, wie erwähnt, zugleich mit einem sekundären Rahmen hergestellt wird. Als die Spinne die Fäden 24 und 26 hergestellt hatte, hatte sie sich dem bereits vorhandenen Faden a (welcher der erste Radialfaden dieses Netzes überhaupt war) soweit genähert, daß in den beiden verbleibenden Winkeln (15° und 17°) kein Platz für weitere Radialfäden blieb. Solche Winkel bezeichnen wir als Restsektoren. Frühere Untersuchungen (Peters 1938) hatten gezeigt, daß die Restsektoren in der Regel kleiner sind als das Doppelte des für den betreffenden Netzbereich geltenden Winkelbetrages. Die Winkelgröße selbst ist nicht ein für allemal festgelegt, sondern von verschiedenen Bedingungen abhängig (Peters 1937). Unter sonst gleichen Umständen befestigt ein und dieselbe Spinne z. B. in einem großen Netz die Radialfäden in einem kleineren Winkelabstand als in einem kleinen. Auch sind die Winkelabstände im oberen Netzteil größer als im unteren. Es ist eine Arbeit im Gang, welche die „Standardmasse“ genau festlegt; sie sind, gleiche Grundbedingungen vorausgesetzt, ziemlich konstant.
In dem angeführten Beispiel gilt für den untersten Netzbereich eine Winkelgröße von 9—10°. Die beiden Restsektoren bei a sind mit 15° bzw. 17° kleiner als das Doppelte dieses Betrages. Ebenso sind z. B. die Sektoren, die an Radialfaden 23 oder 27 anschließen und durch ihre Größe aus der Reihe fallen, Restsektoren. Würde die Spinne, unter Befolgung der eben beschriebenen Grundregel des Speichenziehens, in einen Restsektor noch einen weiteren Radialfaden einfügen, so ergäbe sich ein extrem kleiner Sektor, z. B. links anschließend an Radialfaden a
42
Hans M. Peters, Peter N. Witt und Dieter Wolei
ein solcher von 5—6°. Aber dazu kommt es nicht1, weil die Spinne mit dem Speichenziehen aufhört, sobald sie sich mit ihren Radien einer schon vorhandenen Speiche bis auf einen Winkelabstand genähert hat, der, wie gesagt, kleiner ist als das Doppelte der für den betreffenden Netzbereich geltenden Größe. Bevor die Spinne einen neuen Radial-faden einzieht, greift sie im Zentrum des Netzes, in Rundgängen, die bereits vorhandenen Speichen mit den Beinen ab. Entdeckt sie irgendwo eine Lücke, die noch ausgefüllt werden muß, dann begibt sie sich nach dem Rahmen und zieht einen weiteren Radius ein, bis schließlich, wie gesagt, nur noch Restsektoren vorhanden sind, in die keine Speiche in normalem Abstand jnehr hineinpaßt.
In ganz seltenen Fällen allerdings unterläßt die Spinne das Einziehen eines Radialfadens in eine Lücke, in welche noch eine Speiche hätte eingefügt werden können. Im fertigen Netz stellt sich eine solche Lücke als ein Sektor dar, dessen Winkelgröße das Doppelte der für die betreffende Netzstelle geltenden Winkelgröße übersteigt. Mit vorerst genügender Genauigkeit kann man als die „geltende“ Winkelgröße das Mittel aus den beiden an den großen Sektor rechts und links anschließenden Winkeln annehmen (Weiteres s. unten!). Liegt die Winkelgröße eines Sektors über dem Doppelten der Größe seiner beiden Nachbarsektoren, so definieren wir ihn als „übergroßen Sektor“. Sehr wahrscheinlich handelt es sich dabei stets um übergroße Restsektoren im Sinne der obigen Beschreibung. Das bedarf aber noch weiterer Untersuchung. Wenn ein Sektor durch seine Größe beträchtlich aus der Reihe fällt, so könnte das manchmal auch daran liegen, daß die Spinne an einer Netzstelle plötzlich ein zu großes Maß anwendet. Ein so zustande kommender anomal großer Sektor wäre natürlich kein echter Restsektor.
Wir konnten 263 normale Zilla-Netze auf das Vorkommen übergroßer Sektoren untersuchen. Diese Netze stammten von 45 Individuen verschiedener Altersklassen. 62 Netze waren von Spinnen des vorletzten und letzten Häutungsstadiums hergestellt worden, derselben Altersklassen, auf die sich die gleich zu schildernden Experimente beziehen. Nur 3 von den 263 Netzen enthielten, und zwar je einen übergroßen Sektor (Tabelle 1).
Überraschenderweise stieg nun die Zahl der übergroßen Sektoren in Netzen, welche unter Substanzwirkung hergestellt wurden (Veronal, Morphin, Cardiazol, Coffein, Pervitin, Thyroxin), ganz erheblich an. Dies besagt, daß die Spinnen unter Substanzwirkung häufig Lücken zwischen zwei Radialfäden offen lassen, in welche unbeeinflußte Tiere noch einen Radialfaden eingefügt hätten.
Es konnten 83 Netze untersucht werden, die nach Gaben der eben genannten Substanzen hergestellt worden waren. 22 von ihnen, von
1 Oder vielleicht nur ausnahmsweise 1
9 verschiedenen Tieren gebaut, wiesen zusammen 25 übergroße Sektoren auf1. Der Unterschied gegenüber Normalnetzen ist also, auffallend (26% gegen 1%).
Verfahren der Messung und Berechnung.
Abb. 2 kann zur Erläuterung des Berechnungsverfahrens dienen. Es ergibt sich z. B. für Sektor 13 die Winkelgröße a = 7,5°, für Sektor 15 c = 8,5°. Die Größe des eingeschlossenen Winkels ist b = 19,5°. Da b — (a + c) > 0 (nämlich 3,5°), ist Sektor 14 ein „übergroßer“. Weisen die beiden Anschlußwinkel, wie in diesem Beispiel, eine nur geringe Größendifferenz auf, so kann man ihr Maß ohne Bedenken als das für die betreffende Netzstelle „gültige“ ansehen. Die Beurteüung ist aber schwierig, wenn die Differenz größer ist. Unter Substanzwirkung scheinen nicht selten die Winkelmaße bei der Befestigung der Radialfäden unsicher zu werden. Stellen wir in einem Substanznetz z. B. eine Winkelreihe a = 17°, b = 28°, c = 9° fest, so wäre nach der eben erläuterten Berechnungsweise der mittlere Sektor b als „übergroß“ aufzufassen [b — (a c) = 2°]. Aber hier ist die Störung so stark, daß man den Fall nicht mehr gut mit dein
1 Über die Auswahl dieser Sektoren s. weiter unten!

44 Hans M. Peters, Peter N. Witt und Dieter Wolfe:
in Vergleioh setzen kann, was wir in einem Normalnetz oder auch in einem weniger gestörten Substanznetz als „übergroße Sektoren“ auffassen. Wir haben daher im gegenwärtigen Zusammenhang nur solche Fälle berücksichtigt und in der Tabelle zusammengestellt, in denen die Größendifferenz der beiden benachbarten Winkel bis zu 30% des Betrages des größeren Winkels ausmachte. Diese Differenz dürfte nach dem derzeitigen Stande unserer Kenntnisse über die normale Netzstruktur nicht weit von der Differenz entfernt sein, bis zu der in Normalnetzen benachbarte Sektoren, sofern sie nicht Restsektoren sind, voneinander abweichen.
Es erscheint uns so gut wie sicher, daß die meisten der übergroßen Sektoren der Substanznetze Restsektoren waren, obwohl wir ihre Entstehung nicht protokolliert haben. Aber selbst wenn das falsch wäre, bleibt als sicheres Ergebnis, daß in den unter Substanzeinfluß Tier gestellten Netzen sehr häufig Winkellücken Vorkommen, in welche eine unbeeinflußte Spinne noch einen Radialfaden eingebaut hätte.
Zur Messung der Winkel muß man selbstverständlich den Zustand des Netzes während der Herstellung der Radialfäden rekonstruieren. Denn später, bei der Anfertigung der Befestigungsumgänge in der Warte und beim Einsetzen der Klebfäden, finden Verlagerungen statt, die in Abzug gebracht werden müssen. Wir haben daher als Scheitelpunkt der Sektoren den Mittelpunkt der Netznabe angenommen und von dort die geraden Verbindungslinien nach den Fixierungspunkten an den Rahmenfäden gezogen. Die Annahme des Scheitelpunktes bleibt zwar mit einer gewissen Unsicherheit behaftet, doch sind die möglichen Abweichungen von den wahren Scheitelpunkten der Sektoren so gering, daß sie vernachlässigt werden dürfen.
Bei dem geschilderten Effekt handelt es sich um den Ausfall von Teilhandlungen, die normalerweise zur Durchführung gelangen. Die Handlungsfolge des Speichenziehens kommt nicht zur Vollendung, sondern sie wird vorzeitig abgebrochen. Bei der Veränderung der Feinstruktur des Netzes nach Pervitingaben hatten wir den Eindruck einer mangelhaften Kontrolle der Motorik durch die Sensorik. Im Falle der übergroßen Sektoren kann man nicht im gleichen Sinne von einer sensorischen Störung sprechen. Sollte die Störung überhaupt im Sensorischen liegen, so muß sie von anderer Art sein als im Falle der unregelmäßigen Klebfäden. Wir haben nämlich eine Reihe von Substanznetzen, in denen die Winkelmaße als solche gegenüber normalen Netzen nicht merklich beeinflußt und die auch sonst durchaus „normal“ erscheinen, bei deren Herstellung die Sinneswahrnehmungen der Spinne also intakt gewesen sein mußten, wo aber dennoch übergroße Sektoren auftraten. Man darf den Befund vielleicht auf eine Minderung der zum Vollzug der letzten Etappe des Speichenziehens benötigten Triebenergie zurückführen. Interessant ist es, daß diese geringfügige Störung in einem Falle noch am 4. Tage nach der Substanzabgabe beobachtet wurde. Erst die genaue Durchmusterung von Substanznetzen, die wir zunächst für „normal“ gehalten hatten, lehrte uns in den übergroßen Sektoren eine Störung kennen, die alle anderen an Feinheit übertrifft.
Da wir nach so verschiedenartigen Substanzen wie Veronal, Morphin, Cardiazol, Coffein, Pervitin und Thyroxin übergroße Sektoren auftreten
Die Beeinflussung des Netzbaues der Spinnen.
45
sahen, handelt es sich Wirkung.
o
gewiß
um eine weitgehend substanz-unspezifische
Zusammenfassung der Ergebnisse.
Es ist möglich, mit neurotropen und anderen Substanzen den Netzbau der Radnetzspinne [Zilla x-notata (= litterata)] zu beeinflussen. Daraus scheint sich einerseits ein Weg zur Erforschung der bei der Herstellung des Spinnennetzes integrierten zentralnervösen Prozesse zu ergeben, andererseits eine Möglichkeit zur Ausarbeitung von Tests für pharmakologisch interessierende Substanzen.
Veronal scheint den Netzbautrieb als Ganzes zu hemmen. Manche Substanzen, darunter Pervitin, führen sehr wahrscheinlich zu einer Verkleinerung des Netzes. Nach Coffeingabe wurde ein Netz erhalten, das nur noch die rohe Grundstruktur wiedergibt, aber alle Regelmäßigkeit im einzelnen vermissen läßt. Pervitin bewirkte u. a. Störungen in der Anordnung der Klebfäden; diese nehmen einen mehr oder weniger unregelmäßigen Verlauf. Die gleiche Substanz, sowie einige andere, führen zu Ausfällen bei der Herstellung der Radialfäden.
Wir betrachten das Spinnennetz als ein besonders günstiges Modell zum Studium der zentralen Grundprozesse komplexer Instinkthandlungen. Die Methode der Verwendung neurotroper Substanzen dürfte sich aber zu gleichem Zweck auch auf andere Wirbellose anwenden lassen.
Literatur.
Baltzer, Fr.: Beiträge zur Sinnesphysiologie und Psychologie der Webe-spinnen. Mitt. naturf. Ges., Bern (1923). — Gerhardt, U., u. A. Kästner:
1 Araneae. In Handbuch der Zoologie von Kükenthal u. Krumbach, Bd. 3.
2. Hälfte. Berlin 1938. — Millot, J.: Sens chimiques et sens visuel chez les Arai-gnées. Ann. Biol. 22 (1946). — Peters, H. M.: Studien am Netz der Kreuzspinne (Aranea diadema). I. Z. Morph.u.Ökol. Tiere 32 (1937). — Studien am Netz der Kreuzspinne (Aranea diadema). IL Z. Morph, u. Ökol. Tiere 33 (1938). — Probleme des Kreuzspinnennetzes. Z. Morph, u. Ökol. Tiere 36 (1939). —1 Das Kreuzspinnennetz und seine Probleme. Naturw. 27 (1939). — Zur Geometrie des Spinnen-Netzes. Z. Naturforsch. 2 b (1947). —■ Grundfragen der Tierpsychologie. Stuttgart 1948. — Peters, H. M., u. P. N. Witt: Die Wirkung von Substanzen auf den Netzbau der Spinnen, Experientia 5 (1949). — Spronk, Fr.: Die Abhängigkeit der Netzbauzeiten der Badnetzspinnen Epeira diademata und Zilla x-notata von verschiedenen Außenbedingungen. Z. vergl. Physiol. 22 (1935). —■ Tilqtjin, A.: Influence de la mue sur le comportement du tissage chez les Araignées. J. Psychol. 35 (1938). —- La Toile géométrique des Araignées. Paris 1942. — WiEHLE, H.: Beiträge zur Kenntnis des Radnetzbaues der Epeiriden, Tetra-gnathiden und Uloboriden. Z. Morph, u. ökol. Tiere 8 (1927). — Beiträge zur Biologie der Araneen, insbesondere des Radnetzbaues. Z. Morph, u. ökol. Tiere 11 (1928).—’Weitere Beiträge zur Biologie der Araneen, insbesondere zur Kenntnis des Radnetzbaues. Z. Morph, u. ökol. Tiere 15 (1929). — Neue Beiträge zur Kenntnis des Fanggewebes der Spinnen aus den Familien Argiopidae, Uloboridae und Theridiidae. Z. Morph, u. ökol. Tiere 22 (1931).
Prof. Dr.
Hans M. Peters, Tübingen, Zoolog.
Institut der Universität.
6
Druck der Universitätsdruckerei H. Stürtz AG., Würzburg