Images Collection









View this article in Search Friendly Plain Text
NOTE: This plain text article interpretation has been digitally created by OCR software to estimate the article text, to help both users and search engines find relevant article content. To read the actual article text, view or download the PDF above.
REPRINTED FROM
VÔLUMË 16 • NUMBER 1 • JANUARY 1971
CONTENTS
THE BIOLOGY OF MENTAL HEALTH AND DISEASE—A SYMPOSIUM
Raymond W. Waggoner and William J. Horvath
Issue Editors
1 Changes in Sensory and Motor Systems during Centrally Elicited Attack
by John P. Flynn, Stephen B. Edwards, and Richard J. Bandler, Jr.
20 Hearing and the Neural Detection of Acoustic Patterns
by F. G. Worden
31 The Physiology of Dreaming by Frederick Snyder
45 Modification of Memory Storage Processes
by James L. McGaugh and Ronald G. Dawson
64 Behavioral Gentics by Gerald E. McClearn
82 Biochemical Factors in Anxiety Neurosis by Ferris N. Pitts, Jr.
92 Some Implications of Conditioning Theory for Problems of Drug Abuse by Abraham Wikler
98 Drugs Alter Web-Building of Spiders by Peter N. Witt
114 About the Authors
Reprinted from Behavioral Science Vol. 16, No. 1, January, 1971 Printed in U.S,A,
DRUGS ALTER WEB-BUILDING OF SPIDERS: A REVIEW AND EVALUATION1
by Peter N. Witt
North Carolina Department of Mental Health, Raleigh
Twenty-two years of investigation of spider-web-building and its sensitivity to drugs has produced insight into this invertebrate behavior pattern and its vulnerability. Most data were collected by measuring and analyzing photographs of webs built under different circumstances; groups of web data were subjected to statistical comparisons. Another approach was through analysis of motion pictures of the construction of orbs, built with or without interference. Drugs (chlorpromazine, diazepam, psilocybin), as well as temperature and light conditions could prevent onset of web-building and pentobarbital sodium could cause end of radius construction before completion. D-amphetamine caused irregular radius and spiral spacing, but showed regular execution of probing movements ; the severity of the disturbance in geometry corresponded to drug concentration in the body. Scopolamine caused wide deviation of spiral spacing distinctly different from amphetamine, while LSD-25 application resulted in unusually regular webs. Size of catching area, length of thread, density of structure, thread thickness, and web weight were varied in different ways through treatment with cholinergic and anticholinergic drugs, tranquilizers, etc. Glandular or central nervous system points of attack for drugs are identified, and disturbed webs regarded as the result of interference at any of several levels which contribute to the integrated pattern. Web-building as a biological test method for identification of pathogenic substances in patients’ body fluids is evaluated.
(T*0
INTRODUCTION
Strangely distorted spider webs were observed one morning in the autumn of 1948; the animals had drunk drug solutions the day before. The professor of zoology at the University of Tubingen had enlisted the help of a young pharmacologist to try to shift web-building to a convenient time. The stimulant drugs which were chosen failed to produce the effect, but the webs were built in a way which seemed distorted beyond the range of variations in the geometric pattern which had been observed up to that time (Peters & Witt, 1949). It took only a few days to prove that the phenomenon was reproducible, i.e., every animal which received amphetamine in the evening built a strange web in the morning. It has become a matter of class demonstration and has been confirmed for various spider species (Witt, Reed, & Peakall, 1968; Mardikar, Srinivasan, & Balwani, 1969). The application of certain
1 This work was supported in many different ways; special thanks to the National Science Foundation which provided grant support throughout the last years. Dr. C. F. Reed kindly read this, manuscript and provided generous advice.
drugs to orb-web building spiders at a certain time before web-building results in construction of drug webs. They are distinctly different from any webs which would have been built by that spider at that particular time without the drug.
After more than 20 years of exploring this phenomenon, one asks the question: Have we got beyond the initial surprise of looking at distorted webs? Have we learned much about the conditions under which such webs are built? Can we interpret some of the changes in terms of changed behavior or altered function of the nervous system? How close is the relationship between a certain type of drug and a pattern of change? Has a new experimental , route been found for studying drug effects on behavior?
The following review of studies of web construction and drug effects, carried out by the author, alone and in cooperation with colleagues and students, attempts to answer such questions. It is the author’s conviction that a field of behavioral investigation has been opened which, while only superficially explored and filled with problems, looks increasingly worthwhile to pursue; it promises
Behavioral Science. Volume 16, 1971
Drugs Alter Web-Building of Spiders
99
to provide new insights into behavior of living beings, and into changes of behavior under the influence of drugs.
DRUGS AND THE WEB AS A RECORD OF BEHAVIOR
All observations were made in the laboratory. Building frequency as well as geometry may not be exactly the same for webs buiM in the laboratory as outdoors. Such diff#v onces, however, are irrelevant to the conclusions drawn from thp experiments inasmuch as only data alp used which comp-Sl the drug web with Scq^trb| web (both webs! built by the same spider on different daÆ or webs built by different spiders on thelame ‘dam Significant differences are established by means of statistical methods, comparing web measurg| in an untreated condition with those obtained HjKr treatment.
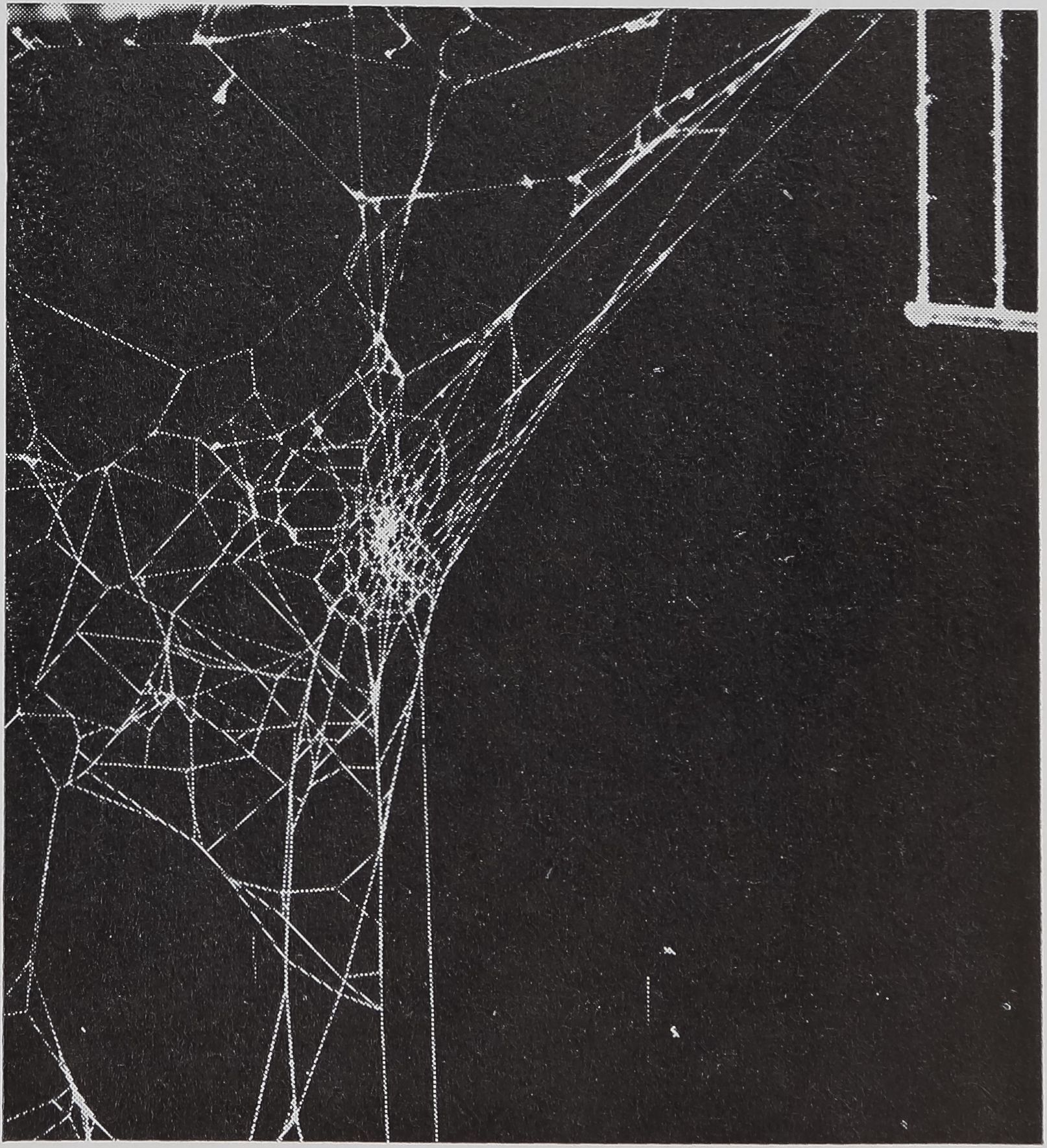
laboratory is maintained at cojffirolled temperature and light conditions, alternating Between an eight hour long cool, dark period and a 16 hour light and warm period. Spiders are kept in individual aluminum fram«| 50 X 50 X 9 cm with removable gl« doors. In earlier «1rs, webs wer@hiokM with ammonium chlorideMcrysHtor sprafed with Krylon white glossy spray paint for photography. Later it w^s found sufficient to place a black box behind the web, fluorescent lightm on all sides, with a screen between web and camera to shield the lens from all light except that reflected by the threads. Photographs with panatomic X film shdpy everyBhread white on a black background and a scale in the upper corner (Fig. lBSftcfed meHESg ments are taken from the re-enlarged film and processed in a computer to generate figures on regularity, size, and shape of the web. A detailed description of the evolution of methods and those used at present in the author’s laboratory can be found in WilBgg al. (1968); this review is concerned predominantly with the work done in that laboratory.
Although early drug experiments had been performed with Zygiella-x-notata CL, an orb-web builder with a free sector in its web, most of the work on drugs and web-building behavior has been carried out on one species of spider, Araneus diadematus Cl., the cross

Routine Bbof.Muy photograph of a web buiff ‘bfpR^adult female Aran^m diadematus CL spider in a flfy X 50 cm aluminum frame. The rigM cornet^ shows the scale – jfthreads 20 mm apart, a lead bar at the bottom, pulling threads verticaMSFo%iiRhe spider in the hub* surrounded by a ring-jgpned free-zone. The sticky Spiral was built from the outside in,[parting at the bottom, and erasing radii in pendulum and round turns to form the ring-shaped catching zone. Radii end peripherally at the frame or split into Y-shaped auxiliaSBframe threads, as seen hefse in Be lo|ker le^Bprner.
spider Rïg. 2jMft’ has been shown that one drug, i.e., d-amphetamine, causes comparable web changes in different species of orb weavers (Witt et ah, 1968). However, there are differen^Sbetween spiders in patterns as well as frequency and regularity of web construction, and || appears advantageous for comparison of results of different experiments to work with one species alone. Where not specifically mentioned, the experiments were carried out with Araneus diadematus Cl., and the pattern of its webs appears in nearly all figures.
Adult female animals, about 4-8 months after hatching from the egg, weigh between 100 and 200 mg and every morning just be-
Behavioral Science, Volume 16, 1971
100
Peter N. Witt

Figv’2. Adult female adematiùsrCl. sitting in ât®isua^^Ki|ing lèStion in ike hub of iH weB|B®ause”»|proper photographie exposure for figg^ip-er, M^fine threads of the web are invisible, Bith ^Méxmption H a stronger threadMjÉfe top. This may beg| pie.ée of the line which wh& laH last a^fên® at the spin,j neret at the rear end of the spider’s^bdonïèn. All 8 legs connect with cephaJ§thora®md are spread in differenfesl diremid#^ moni^Bng vibra^Ri^ along radii. AbsejfMSnHhe 8 e^Hat the rostrMI end of the ..céphalothorax does U.■■ afF^MwS building measurably.
fore sunrise build a web like the one shoyw in Fig. 1. A few more data will illustrate the extent of labor whiclH judged byyhuman standards, appears to be a substantial feat of hard work, precision and beaut* The adult web contains about 0.1 to 0.5 mg of material, i.e. >0.1 percent of the body weighfcbf the builder; this is stretched to a length of 10 to 20 m, fused at every point where thread crosses thread, between 1,000-2,000 times in a web. The animal takes between 20 and 30 minutes to place all those pieces of thread in the right sequence with proper spacing.
After finishing construction, spiders go to the hub of the web, bite out left-over pieces of silk, and stay immobile for hours, head down, the eight legs spread on radii in all directions (Fig. 2). The web is now ready to trap air-borne insects, signal the presence of prey to the spider through thread vibrations, and serviras a path for running to the prey.
The foregoing description has already pointed to the important fact, that the main sensory quality of the spider is that of touch. Even a blinded animal or one building in complete darkness has no difficulty constructing the full-sized web with th^usual regularity in spacing. It is difficult to imagine from human experience, how a world would “look” which was mainly filled with vibrations, tensions, elastic and rigid objects; the existence of an exoskeleton in the invertebrate animal may aid in perceiving the environment that waÿRit provide! ia good reference for deflection of protruding hairs. A close look reveals the legs as rather hairy organs, with bristles standing at right angles to the surface and vibrating in the slightest breeze, other bristles lying nearly flat against the skin, forming short and blunt or long and pointed hairs. Thffe many varieties of hair may represent various specialized pBffptive functions, and though in most ins^piifees the proof is lacking, some micro-Ropic and electrophysiological analysis of hjB»function hiiybeen performed (Foelix, addition,, there exist slit organs of varying size and structure close to the joints, oSr 100 altogether on each animal (Walcott, B969)ralt,‘seems likely that the slit organs become distorted as limbs arSéîretched and bR|> and that tbR provide information on rgative leg and body poMtions. Though the comparison majgseem far fetched, one can imagine a place in the central nervous system of the spider which contains an integrated “Ifuch picture” of the environment similar to the one which may come via our eyes to the visual cortex. Such an assumption is at present not more than a guess derived from behavioral observations, but it is open to electrophysiological and histological testing. We would, therefore, imagine the spider during web construction as an organism which collects information through pulling,
Behavioral Science, Volume 16, 1971
Drugs Alter Web-Building of Spiders
101
probing, stretching its legs, assessing distances, angles and tensions. Such information and stored memory of stages already completed modify the precoded web pattern. The integration of information and program provides the basis for the motions which lead to the positioning of a new piece of thread. And though such words do not make us understand all the neural and biochemical processes which underlie web construction, they help to conceive where and how; a drug can interfere in web building.
In watching construction of the web, it appears like a proceBimiich proceeds with great speed and clockwork precision. As work progre^pflf it becomes increasingly difficult to interrupt the animal ./.^ftipider laying the final turns of the spiral will continue|Æ|h if somebody stepilup close and blows against it. It replaces ;a destroyed thread immediately. Not so at the beginning of web-building. Photographers have jsat\ quiét^Hall night with all their equipment readyMor shooting, only to find that not one of a dozen Bpiders in the room started to build. When they left the laboratory in the middle of the morning, nearly all animals began to build, if the*had reEyed ob
server would return an;hour lSêii to gre^t number of compleSd orb \W|ln the laboratory, the prljdnce of an obSrver Sr pears inhibitory to the start ofBlb-buildinB fclbhe cueSptill unknown. Other confflBpnl in the environment have been identified;/^ supporting or inhibiting web construction. Witt (1956b) observed two groups of two-month-old spiders in a dark room, 71 and 75 individuals, respectively. The first group wap kept at an even temperature, subdivided into smaller groups a|| 15, 20, 20, 24, and 26°C; their w|b-building Sreqlipncy wai decreased 61 percent compared to outdoor controls. The second group went through a daily up and down of temperature equal to the summer day outside; their web-building frequency in the dark was 21 percent higher than that of the other combined groups, but still 40 percent below normal. If light and temperature were both varied according Jb the daily outdoor rhythm, web-building frequency became 97 percent, significantly higher than either of the other groups.
Building occurred at the temperature minimum, just when dark changed to light, before sunrise. Rhythmic light and temperature changes seemed to influence web-building frequency favorably.
Surprisingly enough, spiders have built webs at normal or close to normal frequency under the influence of drugs which severely affected their ability to perform. Fig. 3 shows m web built a||er |§ high dose of d-ampheta-mine; the animal twitched and struggled, but still managed to produce a web of sorts, âo far, three drugs have selectively affected the frequency of web-building: chlorproma-pi||e, diazepam Mgjpsilocybin. Chloroproma-zinfejj (LargafclM (WlfeB 1955) interrupted w*eb-building in 11, 11, pid 21 animals
HBording tp^;l^|Jfcr 1, 2, or 3 dayfi. The highest dos% about 1 /Æmni^àl decreased web-building frequency to 10 percent of controls on three day^after drug application, while the lower doje^ of 0.1 and 0.01 gg/

Fig. 3. An adult female Araneus diadematus Cl. spider weighing 157.9 mg built this structure 12 hours after it had received the relatively high dose of 1 mgS)r about 6.7 g/kg) d-amphetamine in sugar water. Note the remnants of a hub, few irregular and frequently interrupted radii, and some erratic strands of sticky spiral; 24 hours later, this spider built a small and irregular but closer to normal web. The animal’s web structure had recovered completely several days later.
Behavioral Science, Volume 16, 1971
102
Peter N. Witt
animal, respectively, permitted 50 percent of the experimental animals to build webs again on the second day. Nearly as many as in the control group built on the third day after these doses. The number of observations was relatively small, but the graph in the original report suggests a dose-response relationship: In the majority of spiders, the higher the dose of the tranquilizer, the more days without web-building animals. An interesting effect of another tranquilizing drug, diazepam (Valium), will be discussed latir. High doses of psilocybin abolished web-building; lower doses, however, had a characteristic pattern effect which is discussed later.
In contrast to later phases, the ; start of web-building strikes the observer as variable, with many threads being laid only provisionally. Several horizontal threads may be laid nearly parallel to each other; a few can break and are replaced; existingrlhreads are disconnected and shifted to a new position. Pauses of several minutes may occur, fflnder suitable lighting conditions (against a dark background) the observer becomes suddenly aware that a two dimensional structure has evolved, showing the position of the future hub, several radii, and a few frame threads (Fig. 4). It is at this stage that clock-like precision of quick movements appears, and it becomes difficult to interrupt construction.
It is interesting to speculate about the processes which guide the animal at this time. The spider has determined the “size of the future web through the first frame threads, and it has committed itself to a certain density of the mesh structure, at least as far as the radii are concerned, by putting in several of those central angles which will no longer be subdivided later. The silk glands will be empty when the web is finished. The length of thread has already been determined by total available material and body weight of the spider—which requires that thickness of thread be adjusted to it. Taking all this into consideration, one has to conclude that laying out of this part of the web structure requires integration of several different pieces of information. It will become increasingly clear in the course of this discussion, that various

Fig. 4. This photograph was taken in an early stage, while the web was still under construction. Part of frame, hub and radius structure has been completed. Note relatively small radial angles at right and left, which will not be further subdivided,-Tn contrast to wide angles at top and bottom. At this point it is believed that size of future catching area and mesh width have been determined, and that completion of the structure will coincide with exhaustion of silk in glands.
contingencies regulate web proportions. At this point it is essential to note that a detailed plan in pattern and size is already in existence.
There appears a strong tendency to fill the circumference of the web with relatively evenly Spaced radii, the upper ones being usually shorter and bordering wider angles than the lower ones. It has struck observers that subsequent radii are constructed nearly opposite each other, a right horizontal following a left horizontal one, an upper one following a lower one, etc. However, Reed (1969) has shown that the sequence is not invariably determined. By burning radii and observing the position of their replacement, he could show that several sectors of the web may be equivalent in the sense that they possess features capable of eliciting radius
Behavioral Science, Volume 16, 1971
Drugs Alter Web-Building of Spiders
103
building at a given moment. He concludes that sensory guidance in construction of radii may depend upon response to a complex array of forces in the web.
There are a number of radii which tend to be placed late in construction. These require building of an auxiliary frame thread. Without such a structure, they would form a steep angle close to one of the corners of the frame, which could exert strong unidirectional tension on the frame and lead to rupturSApplication of pentobarbital sodium (Nembutal), about 40 jitg/animal, increased significantly the number of oversized sectors, indicating that the drug at this dose did prevent the spider from putting in the late radii (Witt, 1954). This observation has never been followed up with other experiments, and therefore, it appears premature to speculate what kind of change was brought about in the spider to cause such a result.
With each radius completed, the spider returns to the hub and adds a few very short threads. After the last radius has been laid, a prolonged period of hub-adjusting ensues. A new structure emerges, which, in contrast to all other web components, it taken down again before the web is completed. One can observe the rapid construction of a few spiral turns, starting from the hub and ending close to the frame. This has been called the auxiliary or temporary spiral, indicating its nature as a transient structure. It flends strength to the long, elastic radii while the relatively heavy animal lays the narrowly spaced turns of sticky spiral from the periphery inward (Fig. 5).
The temporary nature of the auxiliary spiral has prevented its measurement and its use in drug testing. However, the completely different set of movements which is used to lay this spiral as compared with the movements used in radius construction, has led to the assumption that this is a new phase of web-building (Koenig, 1951). This shift from one set of activities to another also has been used to analyze the problem of the kind of cues which the spider uses in laying the thread. Mere observation of the regular structure and the quick execution of webbuilding would suggest that the process, once set in motion, moves along like the un-

Fig. 5. This spider was interrupted at a later stage of web const ruction than the one in Fig. 4— hub, radii, frame, and auxiliary spiral have been completed. While the sticky spiral was under construction from the periphery inward, the auxiliary spiral was taken down, piecedpy piece. The builder an adulHfemale of Argiope aÆ&ntia Lucas, the golïdÿ|Mgarden wider, orb web builder of a
different genus, but a similar behavior as Araneus diadematus Clerck.
winding of a ^clockwork. This analogy is probably too mechanical; the spider, after all, must adjust to many changes in the environment.
Among other investigators, Peters (1968) tried experimentally to interrupt the spider during spiral building. After 15 minutes in a vial, the spider was returned either to its web as it had left it, to its web with a few of the last laid threads removed, or to the web of another spider which had been interrupted at an earlier or later moment in construction. Three possibilities could be anticipated: (1) that the animal could continue running off the web-building program in the central nervous system, and would therefore take up building at a 15 minute later point in the program, on any structure it encounters; (2) that the animal had stopped the program at the moment of interruption and reactivated
Behavioral Science, Volume 16, 1971
104
Peter N. Witt
it on return to any of the unfinished webs, independent of fit to the existing structure, and (3) that the animal would take its cu^s from the second structure and try to complete it. Peters observed a variety ofjsplu-tions, but found overwhelming evidence for the interpretation that the primary releaser is the existing thread structure, and that the spider uses it as much as- possible for completion of the full orb. His and many other observations point to the importance; of continuous information gathering by the animal about the existing structure. It is the interruption of probing movements, distortion of sensory information, or non-p^e of information elicited through probing f||fS% could lead to the web distortions observed^ after some drugs.
The sti(Hior catching spiral J.fflaid from the periphery inward in narrow circular and pendulum turns. Its construction required a new material,» glue for holding t|fe préÿ\ This comeil from a new get of glands. The sticky spiral construction consumetehe longest period of time in the 20-30 minujil building period. Fastening^or fusing) of the spiral tlpBéii radius occurs o|p| a thoufftnd tim« in an adult web. While it is laid, the temporary spiral is taken down piece by piece. The sticky spiral remains B a record of the movements of the spider. The observer || web constructiorÊotices a set of movements distinctly different from those used previously.. Each leg or pair of legs seems to assume, a special function: The first pair|| predominantly fused to probeBor existing structures like the BBS and next radius, and the last spiral turn; the la^t pair of:legs pullspticky silk from the spinnerets and helps hold; the radius when the new threJS is priRÉ| against it, and the two middle paiBfï legs appear mainly concerned with moving the body and holding it in the right position for thread-laying. The sequence of probing, promoting, and positioning movements’ is peated quickly, over and over again. It should be particularly vulnerable to drug interference, leaving a record in a disorderly spiral.
Interference with regular laying of the sticky spiral in fact is the most striking effect of d-amphetamine (Dexedrine) (Figs. 3 and
6). Amphetamine effects have been reported many times (Peters & Witt|ll949; Peters, Witt, & Wolff, 1950; Wolff & Hempel, 1951; Witt, 1956a; Witt, Brettschneider, & Boris, 1961; Witt et ah, 1968). This well-documented effect is m good example for demonstrating whéther alteration of webbuilding can answer any such pharmacological questions as: (1) is there a drug concentration-effect relationship, Le., does the amount of drug in the body determine the extent of the effect, and (2) can we analyze the mode of action beyond the observation that the spider wafedisturbed?
% FigsJ 3 and 6 make it evident that we can have a more or less disturbed web. While Fig. 6 shows a zigzagging spiral laid across

Fig. 6. An adult; female ArcCnew’s diadematus spider sits in the middliSof its web which it con-Eructed 6 hours after receiving 70 mg/kg dextroamphetamine solution by mouth. Note irregular spacing of radii and spiral turns in comparison to wefcBn Fig. 1, and relatively well preserved web geometry in comparison to Fig. 3, which had been built by another spider after a higher dose of the same drug.
Behavioral Science, Volume 16, 1971
Drugs Alter Web-Building of Spiders
105
slightly irregular central angles, Fig. 3 shows only the remnants of an orb, where the spiral was constructed on an obviously severely disturbed radial structure. Wolff and Hempel (1951) observed that increasing the dose of their amphetamine compound .decreased web-building frequencigin the animals from 20.8 percent after 5-10 /xg to 64.7 percent decrease after 18^-33 /xg per animal. Depending on dose, the effect on central angles can last up to four days after drug application. In a series of experiments, Witt et al. (1961) delayed the disappearance of d-amphetamine from the body of spiders by pretreatment with 600 mg/kg iproniazid, a drug which when used alone had no apparent effect on web-building. If the animal had received 60 mg/kg d-amphetamine alone, the authors could find only 20 percent of the drug left in the body 20 hours later; in contrast, after iproniazid pretreatment, close to 70 Bgfcent of d-amphetamine could be found in the body after 20 hours. The severity of behavioral effects^ were in keeping with the concentration: 60 mg/kg d-amphetamine showed no measurable effects on the webs, but 600 mg/j kg d-amphetamine, as well as 60 mg/kg d-amphetamine after pretreatment with iproniazid, showed changes in size, regularity of central angles, and spiral spacing. The combined effect lasted longer than the single, paralleling rate of d-amphetamine disappearance after pretreatment. The second drug slowed down the catabolism of the first and produced an increased and prolonged effect on web-building. Changes of web-measurement thus proved to be good indicators of drug concentrations in the body.
While the cues which the spider uses for exact placement of radii are unknown, one can observe, particularly in slow motion films, how the animal goes about laying the spiral. Placing of a thread is always preceded by probing with one of the first legs toward the preceding spiral turn. Elimination of the preceding spiral causes searching movements and, finally, deviation of the new spiral toward the second but last turn, which appears now to be the guideline. A frame by frame analysis of motion pictures is useful in de
termining whether the d-amphetamine interferes with the probing movements.
The first film, made by Peters in 1950, revealed that probing with first legs was carried out after d-amphetamine as fre-quenlMas without the drug (Peters et ah, 1950). The spider maintained the normal course of spiral-building, i.e., moving very gradualljpj in circles and pendulum turns from periphery of the web to the center. It alwafl maintained contact with the preceding spiral turn. No jerking movements suggestive of cramp##or severe motor disturbances could be deteSped in the animal. It can be concluded that the block after d-amphetamine occurred between the information collection and the motor execution somewhere in the course of information processing. The block dops not appear to affect either end of the process. Our lack of knowledge, about central nervous^! system proceili! prohibits a more detailed analysis at this time.
lftwf|§ pointed out that the amphetamine treated spider, when laying the spiral, climbs along from radius to radius without losing touch with the preceding spiral turn. Another drug appears to interrupt this process completely. Wolff and Hempel (1951) observed after 30-40 /xg scopolamine per spideritâ loss of direction in spiral construction, so that sometimes at least three partial spirals turned around three centers in different parts of the web. This difference in consequences indicates another important pharmacological fact: Different drugs disturb webs in different ways.
Irregularity of spiral spacing under drugs is a deviation from a norm only slightly more irregular than the control; the spiral at best is far from a perfect geometric structure. One can think of the web as a compromise between regularity, which serves to cover a wide area with a trap of a certain mesh size, and expediency, which demands quick construction. The balance between these two contingencies could be upset in webs built after relatively low doses of d-lysergic acid dietheylamide (LSD-25). Witt (1951) reported that spiders which had received less than 0.05 /xg LSD-25, built webs of larger than control size, with fewer oversized
Behavioral Science, Volume 16, 1971
106
Peter N. Witt
angles and significantly more-regular central angles than in webs built the day before (Fig. 7). The spiral was unusually regularly spaced. However, after higher doses (>0.05 jug), webs were smaller and significantly more irregular.
Building of the sticky spiral in the orb web has shown great sensitivity to drugs. In summary, three drugs have been described as altering the spiral structure in three different ways. The alterations appeared to be correlated to drug concentration in the body/ The form of change permitted tentative interpretation of effects in terms of mode of action and area of attack of The drug. Calibrated web photography provided quantifiable documentation.

Fig. 7. This web is characteristic for Re effect of low doses of d-lysergic acid diethylamide (LSD-25). I| was built by an adult female Zygiella-x-notata spider (note the characteristic free sector, divided by the signal thread), 12 hours after it had received less than 0.05 /xg LSD-25 by mouth in sugar water. Central angles as well as spiral turns were unusually regularly spaced in this as in all other LSD webs.
DRUGS, WEB-SIZE AND WEIGHT
Size of the orb web can be measured in several ways :
a. Length of thread is a rough measure for the distance which the animal covered during construction;
b. Size of catching area, i.e., the ring shaped zone which is covered by the sticky spiral, measures essentially the size of the trap for flying prey;
c. A combination of the two, mesh size, is an expression of the density of the trap or i||J efficiency in catching small prey; it is calculated from the size of the catching area divided by the number of radii and number of spiral turns,
d. Web weight depends on length and thickness of thread, and provides a measure fpr the amount of material which was used for building. The weight is usually determined by measuring the nitrogen content of a web digest, under the assumption that the web polypeptide contains a constant percentage of nitrogen (Witt et al., 1968).
Changes in each of these measures can be produced experimentally. Size changes which are drug independent will be discussed first, in order to interpret the effects under drugs.
A size change most easily observed in the orb web of a spider is that which occurs with age. Fig. 8 shows the early web of a baby spider; it is a small scale, but perfect, image of the adult web in Fig. 1. Witt and Baum (1960) measured size changes in webs of 14 individual Araneus diadematus Cl. during six summer months in the laboratory. They found a steep increase in the size of the catching area from June to August (from 70 to 440 cm2), when a plateau was reached. Radii and spiral turns revealed a different growth pattern, a steep increase in number during the first two months, and an irregular decrease later. The old female spider (Fig. 9) builds a full sized web with relatively little dÿtàil, i.e., a wide-meshed web. Similar changes were recorded by Reed, Witt, and Scarboro (1969) in another species, Argiope aurantia Lucas. Here, too, the mesh size increased throughout, while radius number decreased. Catching area increased steeply first, then decreased toward the end of the animals’ lives. Parallel developmental curves
Behavioral Science, Volume 16, 1971
Drugs Alter Web-Building of Spiders
107
Fig. 8. The photograph shows an early web built by a baby Aweighing 4.3 mg. The animal had hatched from the egg 41 days earlier and had spent the first weeks running around and laying threads on a silk sheet which «shared wUBmany littermates. After young animals draw apart, they start building these small but . regula^structures,-^■ No® tf|l l|)ikes at bottom of picHre indicating 20 mm, and small frame which ^the anima} selected for this structure.
have been described for other species’ webs and may point to a general rule. Measurement of body development was simultaneously recorded. While body weight increased continuously throughout the lifetime of the animals, leg length increased only until the last moult, which coincided with sexual maturity and reaching of full web size, thereafter legs no longer grew.
In the correlation of body and web measures, leg-length was positively related to size of catching area, while weight showed negative association with thread length and with web detail, if measured from the time at which leg growth had stopped. These cor
relations can be rationalized as follows. The long legged, relatively light, mature spider builds a large web with a long thread and high density. The long legged, relatively heavy old spider builds a large web with a short thread and low density. If the important factor distinguishing the two spider populations in these experiments, the mature and the old, was only one of weight, it should be possible to change size characteristcs of webs of mature spiders to those of old ones by increasing their body weight. Christiansen, Baum, and Witt (1962) reported on such experiments.
The mean thread length of 15 webs of 13 spiders was 15.95 ± 1.69 m. The experimenters increased the body weight by 30
Fig. 9. A littermaffl of the spictèr shown in Figure 8 built this web when 25 weeks old. The animal weighed 281.7 mg at that time. Note scale at upper right, and top of small frame at lower left, which permit ’comparison with webs in Figures 1 and 8. The web is relatively large, wide meshed and shows all the characteristic signs of “old age” in the female spider. Such changes in web efficiency occur in spite of regular and ample feeding in the laboratory.
Behavioral Science, Volume 16, 1971


108
Peter N. Witt
percent by placing a piece of lead on the dorsal surface of each abdomen and fastening it with glue. There was no visible interference with mobility. As controls, 12 spiders received the glue treatment alone. The heavier spiders built webs with a mean thread length of 10.76 ± 0.95 m, significantly shorter than before, while the controls showed no significant change in thread length. Removal of weights returned thread length close to original values over three days, indicating slow reversibility of the process. However, the authors measured an additional parameter of the web, its total weight. Mean thread thicknesj; expressed as /ig protein/m thread was recorded as 11.5 ±
2.1 on the control day, as compared to 22.6 ± 5.4 on the day when weight of spiders had been increased, and had returned to
14.1 ±1.5 three days later, a difference significant beyond the 1 percent probability level. It appears, therefore, that while the spider expends a definite amount of thread protein on a given day, its exact disposition in terms of thread length and thickness varies according to circumstances. The spider could have used a full amount of material for a shorter and thicker thread, being forced into this by the necessity to support its heavy body. But what happens to web size as measured in the catching area while thread thickness and length change?
The web Is a trap to catch airborne food. Without a web the spider is unable to get the necessary supply of material and energy for survival. On the p|Ber hand, web construction requires considerable energy and material supply every day. If food is scarce, the animal could build smaller webs to conserve supply, or it could construct large orbs to catch the small prey that is available. Witt (1968a) carried out a number of food deprivation experiments to answer such questions. Twenty-eight spiders, adult and young, were deprived of food for two days, and their webs were swept from the cages so they could not be ingested. Neither web size nor web proportions changed significantly from previous webs, nor was there a measurable change in the amount of thread produced daily. Nine other spiders were deprived of food for six days without showing
changes in web size measures; however, thread supply dropped from an average of 0.49 mg/day to 0.39 mg/day, and body weight decreased by 12 percent. The animals had conserved energy and material by using thinner thread^ but had produced a full-sized trap. Ten days of food deprivation yielded smaller and wider-meshed webs with an Average of 0.3 mg/day of silk; the animal’s body weight had dropped 49 percent. Further extension of the hunger period caused further decrease in web size and body weight and increase in mesh size, but a web was still built daily. Refeeding resulted in an increase, in body weight without immediate change in webs. This series of experiments— like the body weight, thread thickness, web size relationships discussed above—showed the close interaction of behavioral and biochemical functions in adjustment to an emergency. During the first six dayS of hunger, thread production and body weight had decreased, but webs of large area and thin thread were built. Only in longer periods of food deprivation did webs become smaller and wider-meshed, reflecting a severe decrease in silk synthesis. But some silk production continued even then, obviously at the expense of other body constituents, so that there was high loss in body weight. Silk production appears to have so high a priority that it is kept up while tissues shrink. Breed, Levine, and Witt (1964) found that spiders usually take down old webs piece by piece and eat them, before they build new ones. Peakall (1970) Extended thesp observations by placing “cold” spiders on radioactive webs. When he measured radioactivity of successive webs, he found that silk protein was highly conserved by redigesting silk in the web-building cycle.
Considering that web-size is so closely connected with behavioral as well as biochemical processes, it was questioned whether success or failure in catching prey on the web would influence the web. Reed, Witt, Scarboro, and Peakall (1970) raised littermates (Araneus diadematus Cl.), in four different ways, 18 animals in each group. One group was raised in tubes which prevented web-building and these spiders were fed twice a week in the tubes. A second
Behavioral Science, Volume 16, 1971
Drugs Alter Web-Building of Spiders
109
group was raised in tubes, but Jed daily on webs of other spiders so that they had to use the web as a trap. The third group built webs daily or whenever they wanted, and were fed on these webs twice a week. A fourth group built freely and daily, but were always fed by mouth without the help of the web. After about 20 weeks, all animals were given the opportunity to build webs, and the first as well as later webs were carefully measured. It was found that tube-raising affected web size, but such an effect disappeared during the following seven days of observation. Method of feeding, howevS, ;v showSJf no measurable correlation with size measures. Animals built similar webs irrespective of the experience they had with catching food. We conclude on the basis of this evidence that experience with the web as a trap has no visible influence on web size measure||||| Drugs show various effects- on web size, and the foregoing observations should help to interpret such results. Witt (1963b, 1966) reported thlgweb – enlarging effect of physo-stigmine, and Peakall (1964, 1965) analyzed the eff ect Bit the level of the silk glands. Twenty-three spiders received 1 mg/kg physostigmine in sugar water 12 hours before they built a new web, and 23 control animals received sugar water alone. The first webs of the two groups of animals did not differ significantly; but webs built 36 hours after drug application contained significantly more protein material than controls, about twice as much. The physostigmine webs were large, narrow-meshed, and of normal regularity. A day later, all spiders built similar webs| thil control group and drug group being indistinguishable from each other. Peakall (1964) measured the speed of incorporation of radio-actively labelled alanine, an importâM;t> constituent amino acid off spiderfsalk, in thread pulled six hours later from thpanimal. He found that controls under his experimental conditions had an activity of 123 counts per minute in their silk, while physostigmine treated animals had 290 counts per minute, indicating increased synthesis after the drug. When Peakall (1965) employed radioautography to trace the rate of passage of amino acids through the spider’s body, he found quick disappearance of radioactivity from
the hind intestine and blood of physostig-mine-treated spiders. There was relatively fast (4 hour) appearance of tracers in the epithelium and lumen of the silk glands. Control animals were several hours slower in absorbing and using the labelled substances for new silk synthesis. Such data show that the larger-than-control webs built with relatively high quantities of silk after physostigmine are the result of an increase in silk supply ; the drug probably acts primarily at the glandul^f level speeding up silk syn-tl|ësis, and the behaviorffflffBts are the consequence of the close gland-behavior interaction.
Physostigmine is a cholinergic drug; it potentiates the effects of a|ptylcholine, a substance which is widely distributed throughout the body. It appeared possible that the physostigmine effect on speed of silk synthesis was a general cholinergic effect and could be caused by any drug in this group. Even more interesting, an anticholinergic drug like atropine would exert an opposite effect on silk synthesis in the gland. Peakall (1964, 1965) studied this question by measuring speed of alanine incorporation into thread*® well as! tracing autoradiograph-ically labelled amino acids in the spiders’ bodiê|ji His resulwHhow clearly that other cholinergic substances like carbachol and paraoxon have effects on silk synthesis indistinguishable from physostigmine, while atropine slows down the procegg. Witt (1962) fiad feted the effepts of atropine on web patterns and thread production. He found that 1, 2, or 4 mg/kg atropine sulfate given to 19, 19 or 30 spiders by mouth 12 hours before web-building time, resulted in webfi with wider mesh® (shorter thread^ of normal sized catching area after the two lower doses. Significantly smaller and less regular webs were built with less thread after the highest dose. Four mg/kg atropine also reduced by 50 percent the amount of thread which could be pulled from the spiders, again pointing toward thil gland epithelium as primary point of attack for the drug.
We can only speculate what the physiological role of acetylcholine as stimulator of silk production in the spider may be. Acetylcholine usually plays the role of neurotrans-
Behavioral Science, Volume 16, 1971
110
Peter N. Witt
mitter substance, transmitting a signal which has traveled along the nerve to another cell. The sensitivity of the silk gland to this substance could mean that the gland is a receptor of signals from the central nervous system, integrating its speed of silk synthesis with all other components of web-building : it would play a role in start of web-construction, spacing of thread by the legs, and other functions. Though it would well serve to explain many of our findings, such an hypothesis has to wait for histological and biochemical evidence for substantiation.
Several other drugs can apparently cause spiders to build smaller webs. Using the methods of analysis described above] it should be possible to find out whether their primary point of attack in the spider is at the gland or elsewhere. Witt (1960) described the effects of the hallucinatory drug psilocybin as completely abolishing web-building 12 hours after 6 g/kg; reducing it to 5 percent after 600 mg/kg; and reducing it to 11 percent after 300 mg/kg. Of 46 spiders which had received 150 mg/kg psilocybin, 25 drug-webs were obtained the following day. They contained significantly] fewer spiral turns and fewer radii, and had smaller catching areas than controls; mesh size had increased. This looked so similar to webs of heavier spiders, that the author concluded at that time that the drug caused spiders to behave as if they were heavier. However, more complete information obtained later on drugs which caused smaller webs suggests other interpretations which should be retested.
Two sedative drugs, the tranquilizer diazepam (Valium) (Fig. 10) and the Sedative drug phénobarbital (Luminal) (Fig. 11) were tested on spider web-building, using increasing doses, by Reed et al. (1969). In addition to size of catching area, amount and regularity of detail, thread length and web weight, the amount of thread available in the gland was measured. Both drugs produced smaller, lighter webs which could have been the result of less thread supply, or of not making full use of the stored silk.
It should be explained at this point that the guide line can be rolled out until the gland is exhausted. The silk is fastened to

Fig. 10. After 100 mg/kg diazepam (Valium), a tranquilizer, spiders always built relatively small, regular webs. Other investigations showed that the drug did not affect silk synthesis, making it likely that spiders used less than the full silk supply to build webs under diazepam treatment. Compare size to web in Fig. 1.
the axle of a motor while the animal sits on a firm support. Turning the axle rolls out the structural silk, the thread from the am-pullate gland which serves in building hub, radii and frame of the orb web. Histological evidence indicates the probability that construction of the normal web as well as pulling of thread by motor empties the ampullate glands completely.
Reed and Witt (1967) concluded from the diazepam and phénobarbital experiments, that aside from differences in potency and effects on regularity of webs, both drugs produced a similar significant diminution of the activity level of the spider. This was evidenced by reduced thread length and reduced number of spiral turns, combined with no effect on the amount of material available for the web. The latter conclusion was de-
Behavioral Science, Volume 16, 1971
Drugs Alter Web-Building of Spiders
111

Fig. 11. 100 mg/kg phénobarbital (Luminal) caused spiders 36 hours later to build smaller and irregular webs. The scale on the upper right makes it possible to compare size of this web with control in Fig. 1 and diazepam in Fig. 10. Application of high doses of this sedative drug was followed by disturbances in web regularity, which dis^ tinguishes the barbiturate effect from the tranquilizer (Fig. 10).
rived from measuring the amount of silk which could be pulled from spiders which had received the same drug treatment. It must be assumed that the sedated spiders had silk left in the ampullate glands after they had completed their webs, a circumstance which has never yet been observed by us in undrugged spiders.
Small webs can obviously be the result of less than normal silk supply (atropine), or supply of a shorter but thicker thread (body weight increase), or sedation of the builder.
THE WEB TEST
Several other drugs have been investigated, and have been found to alter web geometry. Among these are the more well-known ones like mescaline, caffeine, strychnine, diethyl ether, carbon monoxide, carbon dioxide, nitrous oxide, and adrenochrome. Most have been given by mouth to the spider in a sugar water solution, using ingestion of preorally digested food as the physiological and, therefore, easiest way to get the drug into the animal. Some drugs can become effective through inhalation, and others can be injected. One can compare relative potency of drugs in men and spiders by .referring to the effective dose per kilogram of body weight. In most instances larger dosage
is required for spiders than men, i.e., the drugs are more potent in man than in spiders. Such comparison, however, seems hardly appropriate when one deals with extreme weight differences like a 100-200 mg spider compared to a 50-100 kg man. With few exceptions (chlorpromazine, pentobarbital sodium, phénobarbital) relative potency of drugs in spiders and men has been found similar i.e., drugs rank in a similar order of potency for both (Witt, 1958). Again, this comparison is of questionable value if one considers the criteria which are used to establish a drug effect in either living being, e.g., unusually regular webs vs. hallucinations after LSD-25. The most important finding, seems to be that so many drugs which affect human behavior at various levels alter web geometry of spiders in measurable ways.
This raises thé” following question, Has a biological method been developed which permits identification of small quantities of central nervous system active substances in body fluids? If spiders react to small amounts of LSD-25 for example, they should react to this or similarly active substances, even if they are mixed with many other chemical components in blood, urine, or cerebrospinal fluid. One can assume that many other components of body fluids are inert for spiders, and then test for such hallucinatory substances in biological materials. Such procedures present many methodological problems. If one looks for pathogenic chemicals in mental patients, one has to consider the problem of proper diagnosis, timing in obtaining the sample (acute hallucinatory states), adequate controls, dietetic factors, and many others. Problems à# well as outcome of such biological testing of patients’ body fluids have been discussed by Witt (1958). The findings until today can be summarized as having shown no characteristic biologically active component in patients as compared to healthy controls, which could be established independently in several labora-toriegi There either are no such compounds or the test is not discriminating or sensitive enough.
Another question is that of the contribution which drug-web experiments have made to our knowledge of invertebrate behavior.
Behavioral Science, Volume 16, 1971
112
Peter N. Witt
Looking back, there seems to be a development discernable. While we started out seeing various effects of drugs on a mysterious behavioral process, we became increasingly able to identify those pieces of behavior which were altered by a drug. This paper makes an effort to point out the extent of such identification. Experiments with central nervous system lesions (Witt, 1969), observation of developmental factors in webbuilding (Reed et al., in press), and comparison of drug effects on learned (Le Guelte, 1969) vs. experience-independent behavior, should further help to describe the components of a behavior pattern, their interrelationships, coding, and sensitivity to chemical interference. Our knowledge seems»o have increased interdependent^ on two levels. As we learn more about the basis of web-building behavior, we have improved its use for testing drugs. At the same time, drug experimentation has provided one method for learning more about the basis of the behavior pattern.
REFERENCES
Breed, A. L., Levine, V. D., Peakall, D. B., & Witt, P. N. The fate of the inf act orb web of the spider Araneus diadematu^ Cl. Behaviour, 1964, 23, 43-60.
Christiansen, A., Baum, R., & Witt, P. N. Changes in Kider webs brougH about||y mescaline, psilocybin and an increase fin body weight. J. Pharmacol., 1962. 136, 31-37,
Foelix, R. Structure and function oCt^rsal sensilfe in the spider Araneus diadematus. J. exp. Zool., 1970, 175, 99-124.
Koenig, M. Beitrâge zur Kenntnis des Netzbaus orbiteler Spinnen. Z. Tierpsychol., 1951, 8, 462-493.
Le Guelte, L. Learning in spiders. AmeimZoolo-gist, 1969, 9, 145-152.
Mardikar, B. R., Srinivasan, S., & Balwani, J. H. Effect of centrally acting drugs on |Bhe web building activity of a new ‘Species of an Indian spider. Indian J. med. Sci., 1969, 23 , 550-558.
Peakall, D. B. Effects of cholinergic and anticholinergic drugs on the synthésis of silk fibroins in spiders. Comp, biochem. Physiol., 1964. 12, 465-470.
Peakall, D. B. Regulation of the synthesis of silk fibroins of spiders al^the glandular level. Comp. Physiol., 1965, 15, 509-516.
Peakall, D. B. Conservation of web proteins in the spider Araneus diadematus. J. exp. Zool., in press, 1970.
Peters, H. M., & Witt, P. N. Die Wirkung von
Substanzen auf den Netzbau der Spinnen. Experientia (Basel), 1949, 5, 161.
Peters, H. M., Witt, P. N., & Wolff, D. Die Beein-flfisung des Netzbaues der Spinnen durch neurotrope Substanzen. Z. vergl. Physiol., 1950, 32, 29-44.
Pp|s, P. J. Orb-web construction: Interaction bpfweB» the ^ spider (Araneus diadematus Cl.) and thread configuration. Master of Seifflce Thesis, N. C. State University at Raleigh, 1968′.’ (Publication in press, 1970).
Reed, C. F. Cues in the web-building process. Amer. Zoologist, 1969, 9, 211-221.
Reed, C. F., & Witt, P. N. Progressive disturbance of spider web geometry caused by two sedative drugs. Physiol. Behav., 1967, 3, 119-124.
Reed,^.®., Witt, P. N., & $carboro, M. B. The orb web during the life of Argiope aurantia (Lucas). Develop. Psychobiol., 1969, 2, 120-129.
Reed HH F., Wi||, P. N., Searboro, M. B., & Peakall, D. B. Experience and the orb web. Develop. Psfchobiol., in press 1970.
WalcbjèHC. A’-spider’s vibration reqeptor: Its anatomy and physiology. SBgg Zoologist, 1969, 9, 133-144.
Witt, P. N. d-Lysergsaure-diaethylamid (LSD-25) im Spinnentest. Experientia (Basel), 1951, 7, 310.
Wi#i P. N. Ein biologischer NaAveH von Adre-nochrom und seine mogliche Anwendung. Helv. physiol, pharmacol. A^cta, 1954, 12, 327-337.
Witt, P. N. Die Wirkung einer einmaligen Gabe von Largactil auf den Netzbau der Spinne Zilla-x-notata. Mschr. psychiat. Neurol.,
1955, 129, 123-128.
Witt, P. N. Vergleich der Wirking von Xylopro-pamin und Pervitin auf den Netzbau der Spinne. Arch. inmPharrmcodyn., 1956, 180, 143-152. (a)
Witt, P. N. Die Wirkung von Substanzen auf den Netzba%, der Spinne als biologischer Test. Berlin-Gôttingen-Heidelberg: Springer,
1956. (%
Witt, P. N. The identification of small quantities of hallucinatory substances in body fluids wilflfRife spider test. In C. F. Reed, I. F. Alexander, & S. S. Tomkins. Psychopathology},pa source book. Cambridge: Harvard University Press, 1958.
Witt, P. N. Effects of psilocybin on web-building behavior of spiders. Pharmacologist, 1960, 2, 2.
Witt, P. N. Effects of atropine on spiders’ webbuilding behavior and thread production. Fed. Pw$T, 1962, 21, 2.
Witt, P. N. Environment in relation to the behavior of spideiB Arch, environm. Hlth., 1963,7, 4-12. (a)
Witt, P. N. Interrelationships between web-building behavior and amounts of thread material in the spider Araneus diadematus
Behavioral Science, Volume 16, 1971
Drugs Alter Web-Building of Spiders
113
Cl. Proceed. 16th internat, congr. Zool., 1963, 2, 7. (b)
Witt, P. N. Spider glands and psychotropics.
Mem. inst. Butantan, 1966, 33, 639-644.
Witt, P. N. Behavioral consequences of laser lesions in the central nervous system of Araneus diodematus Cl. Amer. Zoologist, 1969, 9, 121-131.
Witt, P. N., & Baum, R. Changes in orb webs of spiders during growth (Araneus diodematus Clerck and Neoscona vertebrata McCook). Behaviour, 1960, 16, 309-318.
Witt, P. N., Brettschneider, L., & Boris, A. P. Sensitivity to d-amphetamine in spiders after iproniazid and imipramine. J. Pharmacol., 1961, 132, 183-192.
WitB P. N,, Reed, C. F., & Peakall, D. B. A spider’s web. Problems in regulatory biology. New York: Springer Verlag, 1968.
Wolff, D., & Hempel, TJ. Versuche über die Beein-flussung des Netzbaues von Zilla-x-notata durch Pervitin, Scopolamin und Strychnin. Z. vergl. Physiol., 1951, 33, 497-528.
(T’fO
All existences, as long as they retain their character, produce— about themselves, from their essence, in virtue of the power which must be in them—some necessary, outward-facing hypostasis continuously attached to them and representing in image the engendering archetypes: thus fire gives out its heat; snow is cold not merely to itself; fragrant substances are a notable instance; for, as long as they last, something is diffused from them and perceived where-ever they are present.
Plotinus, The Enneads
Behavioral Science, Volume 16, 1971