Images Collection





Read OCR Digitized Article Text
NOTE: This plain text article interpretation has been digitally created by OCR software to estimate the article text, to help both users and search engines find relevant article content. To read the actual article text, view or download the PDF above.
Maturation and d-Amphetamine-Induced Changes in Web Building
CHARLES F. REED Department of Psychology Temple University Philadelphia, Pennsylvania
PETER N. WITT MABEL B. SCARBORO North Carolina Mental Health Research Dorothea Dix Hospital Raleigh, North Carolina
Webs of Araneus diadematus Cl. were obtained under drugged (dextroamphetamine) and control conditions at three age-periods in the development of the spider from juvenility to sexual maturity. Although certain features of construction are affected in all periods and some exhibit differential effects with age, it was not possible to separate the latter from changes in body mass between periods.
The features of the geometrically regular webs are measurably modified when built by spiders drugged with substances which also have behavioral effects in human beings (Witt, 1956; Witt, Reed, & Peakall, 1968). This study examines those modifications in the case of one such drug, dextroamphetamine, for indications of a maturational or age-related vulnerability.
It has been observed that behavioral drugs affect young and old animals differently (Aman & Singh, 1980), suggesting that some factor particular to maturation is responsible. While that may be the case, it is not always possible to easily control for fundamental variables such as body mass. Drug dosage proportional to weight does not necessarily produce proportional effects; in order to obtain an equivalent reaction, small animals often require a larger dose than that which would be proportional to weight. Absence of response in a small animal may be a consequence of inadequate amount of the drug. In order to isolate age-related responses which reflect physicochemical change in the maturing animal, it is necessary to consider that the older animal has also become simply larger.
The process of building the orb web, although an innate program of action, is subtly or grossly modified by the physical constraints upon the spider and by the animal’s physiological condition (Witt et al., 1968).
Applying weights to the animal (Christiansen, Baum, & Witt, 1962; Mayer, 1953) or depleting its thread supply by one means or another (Reed, Witt, Scarboro, & Peakall, 1970) yields a web with a smaller-than-normal catching zone but with an undistorted form. On the other hand, a badly disordered web can be obtained simply by removing radial threads as they are built, until the spider—after many replacements—accepts an inadequate scaffolding to attach its dwindling supply of silk (Koenig, 1951; Reed, 1969). None of these procedures interferes with the sensory physiology of the animal; they alter the cues by which construction is guided.
Reprint requests should be sent to Dr. Charles F. Reed, Department of Psychology, Temple University, Philadelphia, Pennsylvania 19122, U.S.A.
Received for publication 20 March 1981 Revised for publication 15 May 1981 Developmental Psychobiology, 15(1) :61—70 (1982)
©1982 by John Wiley & Sons, Inc. CCC 0012-1630/82/010061-10$01.00
62
REED, WITT, AND SCARBORO
The chief method for changing the physiology of the animal has been by means of drugs which produce behavioral effects in human beings as well (Witt, 1956; Witt et ah, 1968). The alterations in the web are the result of interference with physicochemical events comprising sensory, motor and central nervous system functions. That is not to say that the effects may be clearly distinctive from the physical restraints first described; one common consequence of drug ingestion is reduction in size of the web.
Maturation is of course also a process of modification of the physiological condition of the spider. Sexual adulthood brings increased mass of the spider but also change in the features of the web, for instance, increases in size and in the regularity of the spacing of threads. Moreover, as will be shown, magnitude of alteration of web features in response to a drug appears somehow related to the age of the animal.
Method
Female spiders (Araneus diadematus Cl.) from a single cocoon hatched in the laboratory were housed in individual enclosed cages under controlled conditions of environment and feeding (Witt, 1971).
At three different maturity levels, when the animals were 53, 101, and 166 days old, dextroamphetamine was administered at a time which precedes the usual web-building period by 12-15 hr. The drug was prepared by dissolving dextroamphetamine sulphate in a sugar-water solution (made with half a sugar cube and 10 ml distilled water) in a ratio of 5 mg ^amphetamine : 1 ml sugar water. The spiders were weighed on the day of drug administration and given 1 [A of d-amphetamine solution per 10 mg of body weight (100 mg d-amphetamine/kg). They readily accepted fluids from a syringe ; consumption of each drop was easily observed.
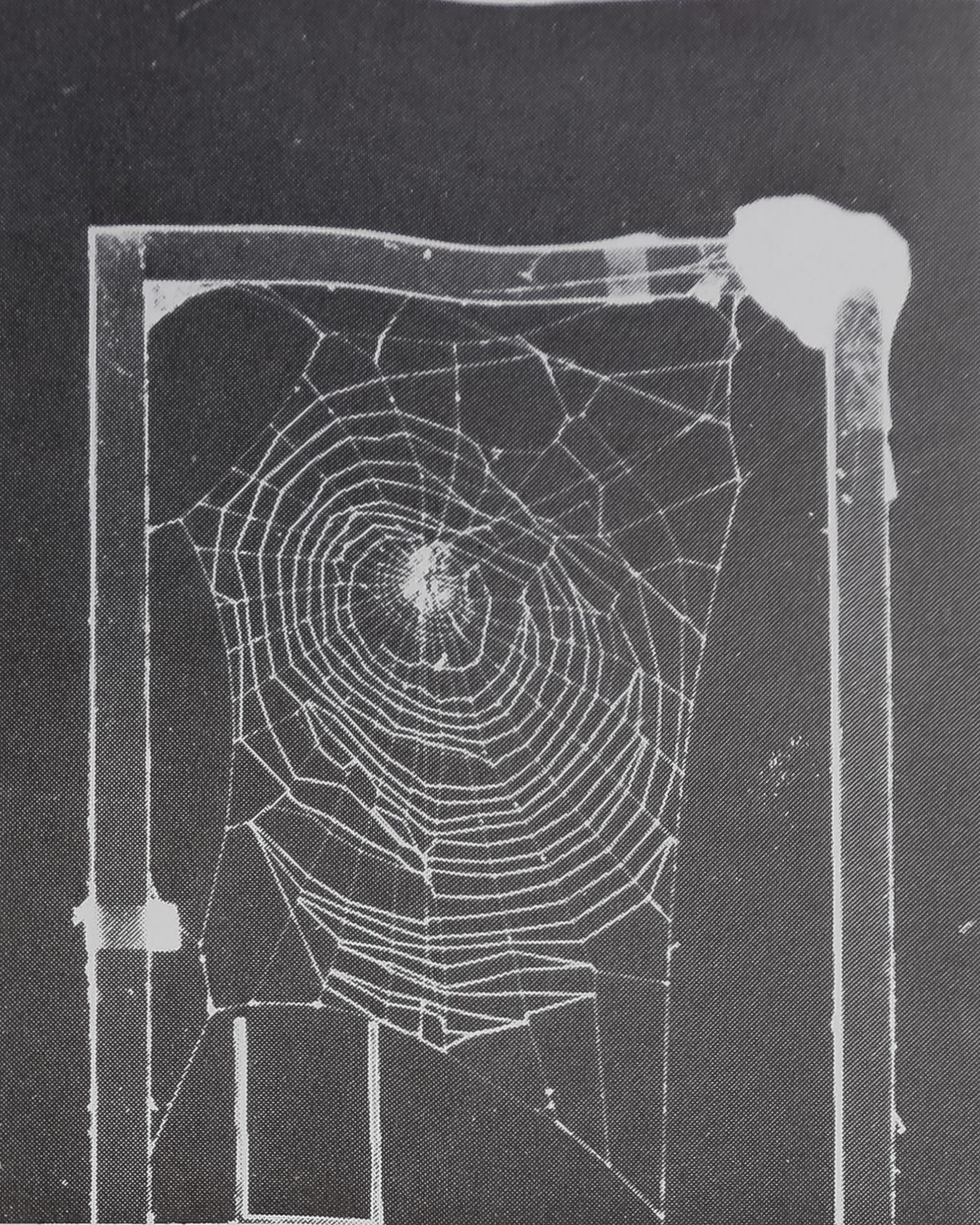
The individual cages are 50 X 50 X 9-cm aluminum, screen, and glass frames which allow ample space for webs built by this species. However, for their webs the youngest spiders, given a choice, more readily adopt a small wooden frame (12 X 16 cm) placed in a larger cage ; it is shown in the first plates in Figure 1.
Each animal served as its own control in each of three maturation periods: a juvenile period at 53 days, a preadult period at 101 days, and an adult period at 166 days following the final molt. Photographs were taken of the web built on the morning of the day of drug administration (control) and of the web built the morning after drug administration. It could not be assured that each animal would build on the designated control and experimental days; complete records, i.e. of control and experimental webs built in each of the three age periods, were obtained for 19 out of 54 animals.1
Immediately after photographing, the webs were collapsed and the remnants left in the cage to be eaten by the spider; this practice insures an entirely new structure for subsequent webs. The material in the old web appears in the ampullate gland 6-24 hr after consumption (Reed et al., 1970).
Web Measurements
An impression of the basic form of the normal web of A. diadematus can be obtained from the upper plates of Figure 1 and from the “control” entries in Table 1.
The “catching zone” of the web is an oval structure elongate vertically with about 30 radii diverging from a hub which is displaced slightly upward (the upmost, North, radius is about 3/7 of the vertical dimension of the catching zone). These features, and the relative uniformity of spacing of the spiral turns which comprise the catching zone,
MATURATION AND ^-AMPHETAMINE WEBS
63
(b)
Fig. 1. Sample control and drug webs at three age periods, (a) and (a’) At 53 days. Constructions of spider (429) the morning before and the morning after ingestion of dextroamphetamine. The small wooden frame preferred by young animals is shown; length calibration is provided by the rod-spacer on the bottom of the cage, (b) and (b’) Same for spider (423) at 101 days. Figure in upper right is calibrating weight, (c) and (c#) Same for spider (423) at 166 days.
64
REED, WITT, AND SCARBORO
Fig. 1. {continued from previous page)
remain stable from immaturity through adulthood. The size of the web—the area of the spiral zone—and the regularity of placement of radial threads increase over the same time span.
By measuring the position of thread-intersections of the representative set of points in the web, its essential characteristics can be reconstructed by computer in a form which
MATURATION AND d-AMPHETAMINE WEBS
65
Fig. 1. {continued from previous page)
permits statistical treatment (Witt,-et al., 1968). Relatively independent measures can be found for the categories size, shape, density, regularity, and element number; a representative set is found in liable 1.
Some of the expressions are self-evident; thread elements refer to the number of radii and number of spiral turns. Shape is expressed in the ratio of vertical and horizontal dimensions of the catching zone. Thread length is the total sum of all thread elements.
Mesh width is the mean area of the compartments into which the spiral zone is divided by the radial and spiral threads. In addition, samples are taken of mesh area in three sectors in the upper (North) and three sectors in the lower (South) portion of the web; the median mesh size is calculated for each portion.
The radial regularity measure . oversized angle, denotes the number of central angles which are greater than twice the size of the smaller of their two adjacent angles .Angle regularity denotes the mean differences, in degrees, between all adjacent central angles in the web.
Relative deviation of spiral is the standard deviation of differences in spiral spacing as a proportion of the mean of those differences: a perfectly regular spacing would have zero relative deviation.2
Results
General Effects of d-Amphetamine
The fundamental features of the web, with representative measures from each, are contained in Table 1. T-tests were performed by the computer as it extracted each measure; the asterisked figures indicate values attaining statistical significance.
There are, first, those web features which were changed by dextroamphetamine at all of the sample periods. The “<i-amphetamine web” is one significantly affected in size of the spiral catching zone, in the number of thread elements (radii and spiral turns), and in
66
REED, WITT, AND SCARBORO
TABLE 1, Web Features in Drug and Control Conditions at Three Maturational Levels.a
| Age | ||||||
| 53 Days | 101 Days | 166 Days | ||||
| Web Feature | Control | Drug | Control | Drug | Control | Drug |
| Thread Elements | ||||||
| Number of radii | 30.4 | 22.7** | 31.0 | 18.1** | 31.5 | 17.8** |
| Number of spirals | 30.2 | **
vd |
42.9 | 13.5** | 56.1 | 15.9** |
| Size | ||||||
| Catching area (cm2 ) | 144 | 58** | 476 | 107** | 749 | 172** |
| Thread length (m) | 10.3 | 3.8** | 22.3 | 4.2** | 32.9 | 6.0** |
| Regularity | ||||||
| Oversize angles (#) | 1.5 | 4.7** | 0.7 | 5.8** | 0.6 | 4.7** |
| Spiral thread (mm) | 0.34 | 0.66** | 0.32 | 0.59** | 0.39 | 0.65** |
| Angle regularity | 3.58 | 10.12** | 2.59 | 11.19** | 2.45 | 12.70** |
| Mesh Density | ||||||
| Mesh width (mm2 ) | 17.1 | 21.5 | 42.3 | 62.7** | 50.6 | 69.5 |
| Upper sample (mm2 ) | 14.6 | 20.3 | 36.1 | 55.5 | 39.6 | 55.8 |
| Lower sample (mm2) | 13.9 | 9.9 | 30.9 | 26.6 | 34.1 | 36.0 |
| Shape | ||||||
| Width/length | 0.79 | 0.92 | 0.81 | 1.01** | 0.86 | 0.99 |
| North/South | 0.76 | 0.66 | 0.74 | 0.64 | 0.74 | 0.68 |
description of^fee web measures is contained in the text. Each entry is the mean value for 19 animals. Significance levels for control-dextroamphetamine comparisons are marked by double asterisks (p < .01).
the precision of placement of threads (regularity). At every age each of these categories differed strongly from control values; the relative magnitude of the change by age will be reviewed below.
Two features of the web, mesh density and shape, showed a less reliable response to the drug than did the first three sets of features. The network of the web, as measured by web-density, was more open—the filter coarser, as it were—in the two older groups as a result of the treatment. However, samples taken close to the vertical axis of the web (“Upper” and “Lower”) gave less evidence of this effect. The lower portion of the web in fact showed an opposite tendency, especially for the 53-day sample. Hence the general loss of density was in other than the lowest sectors of the web.
As regards shape, there was a tendency for loss of the ordinarily oval form of the web in the middle age period; this effect is not easily attributable to reduction of number of spiral turns in the lower web where extra turns normally occur. If spirals were omitted, there should be change in the North/South ratio (the ratio of the upper to the lower radii measuring from the hub); the tables entries indicate no such change.
Relative Magnitude of Change with Age
Since each animal served as its own control in each experiment, the magnitude of change could be expressed as the ratio or as the difference in values in the drugged and control states, and the data analyzed by means of one-way analyses of variance and
MATURATION AND ûf-AMPHETAMINE WEBS
67
covariance. Table 2 contains the means of these ratios, other comparative measures, and the F-ratios obtained in ANOVA and covariance analyses.
The core effects of d-amphetamine (i.e., reductions in thread elements, size of catching zone, and regularity of placement) showed differing degrees of severity of drug effect across age periods. Age differences occurred in responsivity as far as the number of spiral threads, the regularity of radial threads, and the total length of thread were concerned. By contrast, the spiral catching zone appeared to be equally affected in all age periods; its size and its thread regularity were each reduced by dextroamphetamine treatment, but not in degree related to the maturational level of the spider.
Posthoc comparisons (Newman-Keuls) found that in each case in which the ANOVA produced a significant F-ratio, the significant differences involved the youngest age period. The underlined entries in Table 2 are those values for the 101- and 161-day experiments that differed from the 53-day experiments.
For all measures, ^-amphetamine seemed to affect the animals least when they were most immature. It must be recalled, however, that this is a matter of relative severity; the general effect of (i-amphetamine even on the youngest animals was clearly demonstrated. If failure to build a web is taken as a sign of disabling by the drug, evidence of greater rather than lesser sensitivity for the younger animals can be found. In Table 3 are listed the animals which did and did not build webs following drug ingestion. For each age, the
TABLE 2. Relative Change in Web Measures in 3 Age Perils fi
| Age (Days) | F-Ratios^ | ||||
| Web Rature | 53 | 101 | 161 | ANOVA | ANCOVA |
| Thread Elements | |||||
| Number of radii | .77 | .63 | .57 | 3.50 | .28 |
| Number of spirals | .59 | .35 | .29 | 11.26 | 2.20 |
| Size | |||||
| Catching area | .55 | .59 | .23 | 1.55 | 1.04 |
| Thread length | .48 | .25 | .18 | 9.50 | .96 |
| Regularity | |||||
| Oversize angles | 3.21 | 5.05 | 4.11 | 3.29 | 2.58 |
| Spiral thread reg.c | .31 | ,27 | .26 | .12 | .47 |
| Angle regularity Mesh Density | 2.13 | 447 | 4,76 ‘ | 9.36 | 1.75 |
| Mesh width | 1.30 | 1.72, | 1.47 | ‘ 1.02 | 3.95 |
| Upper sample, mesh | 1.91 | 1.73 | 1.27 | .25 | .91; |
| Lower sample, mesh | .93 | IP*09 | 1.01 ‘ | v 1.24 | 1.05 r |
| Shape | |||||
| Width/length | .11 | -.21 | -.14 | 3.51 | 4.30 |
| North/South6 | .09 | .11 | .06 | .12 | .38 |
aEach entry, with the exception of those noted by superscript, represents the ratio of the value of the measure in the ^-amphetamine webs to the value of the measure in the control web. Underlined values indicate significant differences between underlined figures and value in 53-day period (Neumann-Keuls) for the ANOVA.
^For 2 and 53 df, p(.01): 5.03.
cMean deviations for amphetamine minus mean deviations for control (mm); positive: greater regularity in control.
^Control: amphetamine; positive: relative tendency for elongation in drug web. eControl: amphetamine; positive: tendency for upward hub displacement in drug.
68
REED, WITT, AND SCARBORO
TABLE 3. Effect of Dextroamphetamine on Incidence of Web-Building.a
| Age in Days | |||
| 51 | 101 | 161 | |
| Webs after drug | 35 | 38 | 19 |
| No webs after drug | 13 | 1 | 2 |
aBeginning with 54 male and female animals at the first experiment, control webs were required at preselected dates. Columnar sums represent the total number of animals building on each control date; individual entries indicate the number of animals building and not building the next day following drug administration. Mortality and low building frequency by adult males produced an attrition in candidates to 43 and 35 spiders in the two later periods, respectively. The 19 spiders (all females) in the last column, with complete web histories, are the subjects of this report.
animals eligible for drug treatment were those who had built a web at the prescribed control period: 48 had done so at 51 days, 39 at 101 days, and 21 at 161 days. The significant departure from the condition of no relationship (Chi-square « 11.36, p < .01) is attributable chiefly to the relatively low rate of web-building following drug ingestion by the youngest sample (which did not differ from the other samples in web-building frequency under nondrug conditions).
The youngest animals were of course also least in weight. Mean weights for each of the age samples were, respectively by age, 23,119, and 220 mg. Because the animals grew at different rates there was an overlap of weights between the 101-day period and each of the two extremes, but in numbers too small to permit the useful comparison of animals which differed by age but not weight. Unfortunately, the obvious statistical procedure, analysis of covariance with weight as the covariant, becomes questionable as the differences between the control or covariant measures become large, as in this case. Nevertheless, these analyses were performed for each of the measures in Table 2, with each animal’s weight at each of the age periods serving as covariant.
The covariance analysis abolished the age effects found in Table 2. That is to say, the drug, on this analysis, would appear to have had the same effect on the animals irrespective of age.
Discussion
At all ages sampled, the effect of dextroamphetamine ingestion is a web of smaller catching area with a reduced complement of radial and spiral threads, and a loss of precision in placement of threads. The length of the path taken by the spider is shortened, and unless the thickness of the thread has increased in the drugged state—as appears to occur when a normal spider is artificially weighted (Christiansen et al., 1962)—there is a reduced expenditure of thread. It should be noted that the drug was given at a time of day when most of the synthesis of silk material would have been completed and the ampullate glands essentially full (Reed et al., 1970).
These results confirm those performed before the extensive inventory by computer was available (Peters, Witt, & Wolff, 1950); the present study also finds two somewhat more variable effects of d-amphetamine ingestion: a highly variable departure, in the 101-day period, from the normally oval shape of the web, and a loosening of mesh density in the two older periods. Interestingly, this loosening or loss of fineness of density does not seem to occur in the lower portion of the web, a position which is favored in catching,
MATURATION AND d-AMPHET AMINE WEBS
69
since the lurking position of the spider is cephalothorax-downward, facing the lower portion of the catching zone.
Because of its smaller size and somewhat open network, the ^-amphetamine web is presumably a less effective snare than is the normal web. Note that the web geometry is not drastically disrupted: all features of the normal web are present in the d-amphetamine web, albeit in altered dimensions. The successive stages of construction—placement of radial threads, positioning of hub, placement of the spiral threads, retention of a free zone near the hub—have been accomplished by the spider, even if done with apparently contracted expenditure of effort and perhaps of material.
Some features of the web program are unaffected by the experimental treatment. Despite a dosage effective in altering web geometry, the process of building the web is initiated and executed. All of the elements of the usual program are present. The general form of the web is recognizable: radii, spirals and hub are distinct, enumerable and generally in place. Higher dosage levels would produce greater disturbance (Witt, 1971) but comparative measurement would become meaningless when semblance of the normal geometry was lost.
Some features, while present, have been dimensionally altered, at least to some degree in all age periods. The linear extent and possibly the amount of thread is less than normal: fewer radii have been placed and fewer spiral turns placed upon them. Hence the area of the web proper is diminished. The filter formed by radial and spiral threads is coarser, in part because the threads have not been placed with customary precision: some measuring function appears to have been disturbed. The degree of spiral zone alteration remains approximately the same at all periods; it is not possible to say whether it is equally disturbed or equally preserved.
Some features, finally, appear relatively immune, in the juvenily period, to disturbances which occur at older periods. Thread length, spiral-number and angle regularity all seem relatively preserved in the juvenile period. It is these differences however which evaporate in the covariance analysis.
The covariance analysis suggests that were weights somehow to be made equal at all ages, the drug effects would also be equivalent, even for those features which appear to differ by age otherwise. The difficulty in that inference of course is that the difference in weights may simply be indices of other critical differences between the several periods of life somehow correlated with weight. It is difficult to counter that argument, although it must be granted that the reasons for small effect in animals of small mass must be explained at some point.
As difficult as it is to identify the bases for age differences in the responses of spiders, a more formidable problem faces identification of the bases of variability in drug response in complex animals, especially human beings. The biochemical processes involved in the action of dextroamphetamine appear to be similar in spiders and in men (Peters et al., 1950), putting aside obvious differences in structural complexity. Web construction is a particularly narrow kind of behavior, regulated by sensory feedback but apparently unaffected by experience (Mayer, 1953; Petrusewiczowa, 1938; Reed et al., 1970). It does have the virtue of providing a rich and natural sample of behavior without encountering the difficulties which an experimenter confronts in choosing suitably representative and adequate behavioral indices for human behavior. Motivational problems such as the disinclination of the drugged human subject to tolerate experimental procedures are not encountered; the building of the web is a paramount requirement for the spider, making it highly likely that it will display any disabling effects of the drug. Unfortunately, it is not easy to identify a behavior which has a similar preferment in human beings. Given the difficulty in identifying independent maturational effects in the rather predictable, rigidly
70
REED, WITT, AND SCARBORO
patterned behavior of building webs, it is necessary to be cautious about drawing comparisons to changes in the complex behaviors of human adolescents and adults.
Notes
1 Requiring a web on each of six particular days, three of which inyolve drug effects, is stringent even for the high rate of building (approaching one web per day) which females of this species show at the period of life spanned in the experiments. (See Table 3.)
2 Loss of precision is more directly interpretable for spiral turns than it is for radii. In the process of attaching the spiral thread, clear probing movements of the first pair of legs can be observed; removing them disturbs the precision of the spiral (Reed, Witt, & Jones, 1965). For our radial precision measures, there is some inevitable apparent loss of precision because the spider does not fill the web space by a successive bisection of unfilled sectors. This spurious loss of precision would become greater as the number of radii fell below normal.
References
Aman, M. Q., and Singh, N. N. (1980). The usefulness of thioridazine for treating childhood disorders-fact or folklore? Amer. J. Ment. Def, 84:331-338.
Christiansen, A., Baum, R., and Witt, P. N. (1962). Changes in spider webs brought about by mescalin, psilocybin, and an increase in body weight. J. Pharm. Exp. Therap, 136:31-31.
Mayer; G. (1953). Untersuchungen ueber Herstellung und Struktur des Radnetzes von Aranea diadema und Zilla-x-notata mit besonderer Berücksichtigung des Unterschieds von Jugend- und Alters-netzen. Z. Tierpsychol. 9:337-362.
Koenig, M. (1951). Beitrage zur Kenntnis des Netzbaus orbiteier Spinnen. Z. Tierpsychol., 5:462-493. Peters, H. M., Witt, P. N., and Wolff, D. (1950). Die Beeinflussung des Netzbaues der Spinnen. Z. Vergl. Physiol., 32:29-44.
Petrusewiczowa, E. (1938). Beobachtungen über den Bau des Netzes der Spinnen. Trav. Inst. Biol.
Univ. Wilna, 9:1-25.
Reed, C. F. (1969). Cues in the web building process. Am. Zool., 9:211-221.
Reed, C. F., Witt, P. N., and Jones, R. L. (1965). The measuring function of the first legs of Araneus diadematus Cl. Behaviour, 25:98-119.
Reed, C. F., Witt, P. N., Scarboro, M. B., and Peakall, D. B. (1970). Experience and the orb web. Dev. Psychobiol, 3:251-265.
Witt, P. N. (1956). Die Wirkung von Substanzen auf den Netzhau der Spinne als Biologischer Test. Berlin: Springer Verlag.
Witt, P. N. (1961). Web building. In P. Gray (ed.), The Encyclopedia of the Biological Sciences. New York: Reinhold. Pp. 1072-1073.
Witt, P. N. (1971a). Drugs alter web-building of spiders. Behav. Sei, 76:98-113.
Witt, P. N. (1971b). Instructions for working with web-building spiders in the laboratory. BioScience, 21:23-25.
Witt, P. N., Reed, C. F., and Peakall, D. B. (1968). A Spider’s Web. Berlin: Springer Verlag,
Webs of Araneus diadematus Cl. were obtained under drugged (dextroamphetamine) and control conditions at three age-periods in the development of the spider from juvenility to sexual maturity. Although certain features of construction are affected in all periods and some exhibit differential effects with age, it was not possible to separate the latter from changes in body mass between periods.
The features of the geometrically regular webs are measurably modified when built by spiders drugged with substances which also have behavioral effects in human beings (Witt, 1956; Witt, Reed, & Peakall, 1968). This study examines those modifications in the case of one such drug, dextroamphetamine, for indications of a maturational or age-related vulnerability.
It has been observed that behavioral drugs affect young and old animals differently (Aman & Singh, 1980), suggesting that some factor particular to maturation is responsible. While that may be the case, it is not always possible to easily control for fundamental variables such as body mass. Drug dosage proportional to weight does not necessarily produce proportional effects; in order to obtain an equivalent reaction, small animals often require a larger dose than that which would be proportional to weight. Absence of response in a small animal may be a consequence of inadequate amount of the drug. In order to isolate age-related responses which reflect physicochemical change in the maturing animal, it is necessary to consider that the older animal has also become simply larger.
The process of building the orb web, although an innate program of action, is subtly or grossly modified by the physical constraints upon the spider and by the animal’s physiological condition (Witt et al., 1968).
Applying weights to the animal (Christiansen, Baum, & Witt, 1962; Mayer, 1953) or depleting its thread supply by one means or another (Reed, Witt, Scarboro, & Peakall, 1970) yields a web with a smaller-than-normal catching zone but with an undistorted form. On the other hand, a badly disordered web can be obtained simply by removing radial threads as they are built, until the spider—after many replacements—accepts an inadequate scaffolding to attach its dwindling supply of silk (Koenig, 1951; Reed, 1969). None of these procedures interferes with the sensory physiology of the animal; they alter the cues by which construction is guided.
Reprint requests should be sent to Dr. Charles F. Reed, Department of Psychology, Temple University, Philadelphia, Pennsylvania 19122, U.S.A.
Received for publication 20 March 1981 Revised for publication 15 May 1981 Developmental Psychobiology, 15(1) :61—70 (1982)
©1982 by John Wiley & Sons, Inc. CCC 0012-1630/82/010061-10$01.00
62
REED, WITT, AND SCARBORO
The chief method for changing the physiology of the animal has been by means of drugs which produce behavioral effects in human beings as well (Witt, 1956; Witt et ah, 1968). The alterations in the web are the result of interference with physicochemical events comprising sensory, motor and central nervous system functions. That is not to say that the effects may be clearly distinctive from the physical restraints first described; one common consequence of drug ingestion is reduction in size of the web.
Maturation is of course also a process of modification of the physiological condition of the spider. Sexual adulthood brings increased mass of the spider but also change in the features of the web, for instance, increases in size and in the regularity of the spacing of threads. Moreover, as will be shown, magnitude of alteration of web features in response to a drug appears somehow related to the age of the animal.
Method
Female spiders (Araneus diadematus Cl.) from a single cocoon hatched in the laboratory were housed in individual enclosed cages under controlled conditions of environment and feeding (Witt, 1971).
At three different maturity levels, when the animals were 53, 101, and 166 days old, dextroamphetamine was administered at a time which precedes the usual web-building period by 12-15 hr. The drug was prepared by dissolving dextroamphetamine sulphate in a sugar-water solution (made with half a sugar cube and 10 ml distilled water) in a ratio of 5 mg ^amphetamine : 1 ml sugar water. The spiders were weighed on the day of drug administration and given 1 [A of d-amphetamine solution per 10 mg of body weight (100 mg d-amphetamine/kg). They readily accepted fluids from a syringe ; consumption of each drop was easily observed.
The individual cages are 50 X 50 X 9-cm aluminum, screen, and glass frames which allow ample space for webs built by this species. However, for their webs the youngest spiders, given a choice, more readily adopt a small wooden frame (12 X 16 cm) placed in a larger cage ; it is shown in the first plates in Figure 1.
Each animal served as its own control in each of three maturation periods: a juvenile period at 53 days, a preadult period at 101 days, and an adult period at 166 days following the final molt. Photographs were taken of the web built on the morning of the day of drug administration (control) and of the web built the morning after drug administration. It could not be assured that each animal would build on the designated control and experimental days; complete records, i.e. of control and experimental webs built in each of the three age periods, were obtained for 19 out of 54 animals.1
Immediately after photographing, the webs were collapsed and the remnants left in the cage to be eaten by the spider; this practice insures an entirely new structure for subsequent webs. The material in the old web appears in the ampullate gland 6-24 hr after consumption (Reed et al., 1970).
Web Measurements
An impression of the basic form of the normal web of A. diadematus can be obtained from the upper plates of Figure 1 and from the “control” entries in Table 1.
The “catching zone” of the web is an oval structure elongate vertically with about 30 radii diverging from a hub which is displaced slightly upward (the upmost, North, radius is about 3/7 of the vertical dimension of the catching zone). These features, and the relative uniformity of spacing of the spiral turns which comprise the catching zone,
MATURATION AND ^-AMPHETAMINE WEBS
63
(b)
Fig. 1. Sample control and drug webs at three age periods, (a) and (a’) At 53 days. Constructions of spider (429) the morning before and the morning after ingestion of dextroamphetamine. The small wooden frame preferred by young animals is shown; length calibration is provided by the rod-spacer on the bottom of the cage, (b) and (b’) Same for spider (423) at 101 days. Figure in upper right is calibrating weight, (c) and (c#) Same for spider (423) at 166 days.



64
REED, WITT, AND SCARBORO

Fig. 1. {continued from previous page)
remain stable from immaturity through adulthood. The size of the web—the area of the spiral zone—and the regularity of placement of radial threads increase over the same time span.
By measuring the position of thread-intersections of the representative set of points in the web, its essential characteristics can be reconstructed by computer in a form which

MATURATION AND d-AMPHETAMINE WEBS
65

Fig. 1. {continued from previous page)
permits statistical treatment (Witt,-et al., 1968). Relatively independent measures can be found for the categories size, shape, density, regularity, and element number; a representative set is found in liable 1.
Some of the expressions are self-evident; thread elements refer to the number of radii and number of spiral turns. Shape is expressed in the ratio of vertical and horizontal dimensions of the catching zone. Thread length is the total sum of all thread elements.
Mesh width is the mean area of the compartments into which the spiral zone is divided by the radial and spiral threads. In addition, samples are taken of mesh area in three sectors in the upper (North) and three sectors in the lower (South) portion of the web; the median mesh size is calculated for each portion.
The radial regularity measure . oversized angle, denotes the number of central angles which are greater than twice the size of the smaller of their two adjacent angles .Angle regularity denotes the mean differences, in degrees, between all adjacent central angles in the web.
Relative deviation of spiral is the standard deviation of differences in spiral spacing as a proportion of the mean of those differences: a perfectly regular spacing would have zero relative deviation.2
Results
General Effects of d-Amphetamine
The fundamental features of the web, with representative measures from each, are contained in Table 1. T-tests were performed by the computer as it extracted each measure; the asterisked figures indicate values attaining statistical significance.
There are, first, those web features which were changed by dextroamphetamine at all of the sample periods. The “<i-amphetamine web” is one significantly affected in size of the spiral catching zone, in the number of thread elements (radii and spiral turns), and in
66
REED, WITT, AND SCARBORO
TABLE 1, Web Features in Drug and Control Conditions at Three Maturational Levels.a
| Age | ||||||
| 53 Days | 101 Days | 166 Days | ||||
| Web Feature | Control | Drug | Control | Drug | Control | Drug |
| Thread Elements | ||||||
| Number of radii | 30.4 | 22.7** | 31.0 | 18.1** | 31.5 | 17.8** |
| Number of spirals | 30.2 | **
vd |
42.9 | 13.5** | 56.1 | 15.9** |
| Size | ||||||
| Catching area (cm2 ) | 144 | 58** | 476 | 107** | 749 | 172** |
| Thread length (m) | 10.3 | 3.8** | 22.3 | 4.2** | 32.9 | 6.0** |
| Regularity | ||||||
| Oversize angles (#) | 1.5 | 4.7** | 0.7 | 5.8** | 0.6 | 4.7** |
| Spiral thread (mm) | 0.34 | 0.66** | 0.32 | 0.59** | 0.39 | 0.65** |
| Angle regularity | 3.58 | 10.12** | 2.59 | 11.19** | 2.45 | 12.70** |
| Mesh Density | ||||||
| Mesh width (mm2 ) | 17.1 | 21.5 | 42.3 | 62.7** | 50.6 | 69.5 |
| Upper sample (mm2 ) | 14.6 | 20.3 | 36.1 | 55.5 | 39.6 | 55.8 |
| Lower sample (mm2) | 13.9 | 9.9 | 30.9 | 26.6 | 34.1 | 36.0 |
| Shape | ||||||
| Width/length | 0.79 | 0.92 | 0.81 | 1.01** | 0.86 | 0.99 |
| North/South | 0.76 | 0.66 | 0.74 | 0.64 | 0.74 | 0.68 |
description of^fee web measures is contained in the text. Each entry is the mean value for 19 animals. Significance levels for control-dextroamphetamine comparisons are marked by double asterisks (p < .01).
the precision of placement of threads (regularity). At every age each of these categories differed strongly from control values; the relative magnitude of the change by age will be reviewed below.
Two features of the web, mesh density and shape, showed a less reliable response to the drug than did the first three sets of features. The network of the web, as measured by web-density, was more open—the filter coarser, as it were—in the two older groups as a result of the treatment. However, samples taken close to the vertical axis of the web (“Upper” and “Lower”) gave less evidence of this effect. The lower portion of the web in fact showed an opposite tendency, especially for the 53-day sample. Hence the general loss of density was in other than the lowest sectors of the web.
As regards shape, there was a tendency for loss of the ordinarily oval form of the web in the middle age period; this effect is not easily attributable to reduction of number of spiral turns in the lower web where extra turns normally occur. If spirals were omitted, there should be change in the North/South ratio (the ratio of the upper to the lower radii measuring from the hub); the tables entries indicate no such change.
Relative Magnitude of Change with Age
Since each animal served as its own control in each experiment, the magnitude of change could be expressed as the ratio or as the difference in values in the drugged and control states, and the data analyzed by means of one-way analyses of variance and
MATURATION AND ûf-AMPHETAMINE WEBS
67
covariance. Table 2 contains the means of these ratios, other comparative measures, and the F-ratios obtained in ANOVA and covariance analyses.
The core effects of d-amphetamine (i.e., reductions in thread elements, size of catching zone, and regularity of placement) showed differing degrees of severity of drug effect across age periods. Age differences occurred in responsivity as far as the number of spiral threads, the regularity of radial threads, and the total length of thread were concerned. By contrast, the spiral catching zone appeared to be equally affected in all age periods; its size and its thread regularity were each reduced by dextroamphetamine treatment, but not in degree related to the maturational level of the spider.
Posthoc comparisons (Newman-Keuls) found that in each case in which the ANOVA produced a significant F-ratio, the significant differences involved the youngest age period. The underlined entries in Table 2 are those values for the 101- and 161-day experiments that differed from the 53-day experiments.
For all measures, ^-amphetamine seemed to affect the animals least when they were most immature. It must be recalled, however, that this is a matter of relative severity; the general effect of (i-amphetamine even on the youngest animals was clearly demonstrated. If failure to build a web is taken as a sign of disabling by the drug, evidence of greater rather than lesser sensitivity for the younger animals can be found. In Table 3 are listed the animals which did and did not build webs following drug ingestion. For each age, the
TABLE 2. Relative Change in Web Measures in 3 Age Perils fi
| Age (Days) | F-Ratios^ | ||||
| Web Rature | 53 | 101 | 161 | ANOVA | ANCOVA |
| Thread Elements | |||||
| Number of radii | .77 | .63 | .57 | 3.50 | .28 |
| Number of spirals | .59 | .35 | .29 | 11.26 | 2.20 |
| Size | |||||
| Catching area | .55 | .59 | .23 | 1.55 | 1.04 |
| Thread length | .48 | .25 | .18 | 9.50 | .96 |
| Regularity | |||||
| Oversize angles | 3.21 | 5.05 | 4.11 | 3.29 | 2.58 |
| Spiral thread reg.c | .31 | ,27 | .26 | .12 | .47 |
| Angle regularity Mesh Density | 2.13 | 447 | 4,76 ‘ | 9.36 | 1.75 |
| Mesh width | 1.30 | 1.72, | 1.47 | ‘ 1.02 | 3.95 |
| Upper sample, mesh | 1.91 | 1.73 | 1.27 | .25 | .91; |
| Lower sample, mesh | .93 | IP*09 | 1.01 ‘ | v 1.24 | 1.05 r |
| Shape | |||||
| Width/length | .11 | -.21 | -.14 | 3.51 | 4.30 |
| North/South6 | .09 | .11 | .06 | .12 | .38 |
aEach entry, with the exception of those noted by superscript, represents the ratio of the value of the measure in the ^-amphetamine webs to the value of the measure in the control web. Underlined values indicate significant differences between underlined figures and value in 53-day period (Neumann-Keuls) for the ANOVA.
^For 2 and 53 df, p(.01): 5.03.
cMean deviations for amphetamine minus mean deviations for control (mm); positive: greater regularity in control.
^Control: amphetamine; positive: relative tendency for elongation in drug web. eControl: amphetamine; positive: tendency for upward hub displacement in drug.
68
REED, WITT, AND SCARBORO
TABLE 3. Effect of Dextroamphetamine on Incidence of Web-Building.a
| Age in Days | |||
| 51 | 101 | 161 | |
| Webs after drug | 35 | 38 | 19 |
| No webs after drug | 13 | 1 | 2 |
aBeginning with 54 male and female animals at the first experiment, control webs were required at preselected dates. Columnar sums represent the total number of animals building on each control date; individual entries indicate the number of animals building and not building the next day following drug administration. Mortality and low building frequency by adult males produced an attrition in candidates to 43 and 35 spiders in the two later periods, respectively. The 19 spiders (all females) in the last column, with complete web histories, are the subjects of this report.
animals eligible for drug treatment were those who had built a web at the prescribed control period: 48 had done so at 51 days, 39 at 101 days, and 21 at 161 days. The significant departure from the condition of no relationship (Chi-square « 11.36, p < .01) is attributable chiefly to the relatively low rate of web-building following drug ingestion by the youngest sample (which did not differ from the other samples in web-building frequency under nondrug conditions).
The youngest animals were of course also least in weight. Mean weights for each of the age samples were, respectively by age, 23,119, and 220 mg. Because the animals grew at different rates there was an overlap of weights between the 101-day period and each of the two extremes, but in numbers too small to permit the useful comparison of animals which differed by age but not weight. Unfortunately, the obvious statistical procedure, analysis of covariance with weight as the covariant, becomes questionable as the differences between the control or covariant measures become large, as in this case. Nevertheless, these analyses were performed for each of the measures in Table 2, with each animal’s weight at each of the age periods serving as covariant.
The covariance analysis abolished the age effects found in Table 2. That is to say, the drug, on this analysis, would appear to have had the same effect on the animals irrespective of age.
Discussion
At all ages sampled, the effect of dextroamphetamine ingestion is a web of smaller catching area with a reduced complement of radial and spiral threads, and a loss of precision in placement of threads. The length of the path taken by the spider is shortened, and unless the thickness of the thread has increased in the drugged state—as appears to occur when a normal spider is artificially weighted (Christiansen et al., 1962)—there is a reduced expenditure of thread. It should be noted that the drug was given at a time of day when most of the synthesis of silk material would have been completed and the ampullate glands essentially full (Reed et al., 1970).
These results confirm those performed before the extensive inventory by computer was available (Peters, Witt, & Wolff, 1950); the present study also finds two somewhat more variable effects of d-amphetamine ingestion: a highly variable departure, in the 101-day period, from the normally oval shape of the web, and a loosening of mesh density in the two older periods. Interestingly, this loosening or loss of fineness of density does not seem to occur in the lower portion of the web, a position which is favored in catching,
MATURATION AND d-AMPHET AMINE WEBS
69
since the lurking position of the spider is cephalothorax-downward, facing the lower portion of the catching zone.
Because of its smaller size and somewhat open network, the ^-amphetamine web is presumably a less effective snare than is the normal web. Note that the web geometry is not drastically disrupted: all features of the normal web are present in the d-amphetamine web, albeit in altered dimensions. The successive stages of construction—placement of radial threads, positioning of hub, placement of the spiral threads, retention of a free zone near the hub—have been accomplished by the spider, even if done with apparently contracted expenditure of effort and perhaps of material.
Some features of the web program are unaffected by the experimental treatment. Despite a dosage effective in altering web geometry, the process of building the web is initiated and executed. All of the elements of the usual program are present. The general form of the web is recognizable: radii, spirals and hub are distinct, enumerable and generally in place. Higher dosage levels would produce greater disturbance (Witt, 1971) but comparative measurement would become meaningless when semblance of the normal geometry was lost.
Some features, while present, have been dimensionally altered, at least to some degree in all age periods. The linear extent and possibly the amount of thread is less than normal: fewer radii have been placed and fewer spiral turns placed upon them. Hence the area of the web proper is diminished. The filter formed by radial and spiral threads is coarser, in part because the threads have not been placed with customary precision: some measuring function appears to have been disturbed. The degree of spiral zone alteration remains approximately the same at all periods; it is not possible to say whether it is equally disturbed or equally preserved.
Some features, finally, appear relatively immune, in the juvenily period, to disturbances which occur at older periods. Thread length, spiral-number and angle regularity all seem relatively preserved in the juvenile period. It is these differences however which evaporate in the covariance analysis.
The covariance analysis suggests that were weights somehow to be made equal at all ages, the drug effects would also be equivalent, even for those features which appear to differ by age otherwise. The difficulty in that inference of course is that the difference in weights may simply be indices of other critical differences between the several periods of life somehow correlated with weight. It is difficult to counter that argument, although it must be granted that the reasons for small effect in animals of small mass must be explained at some point.
As difficult as it is to identify the bases for age differences in the responses of spiders, a more formidable problem faces identification of the bases of variability in drug response in complex animals, especially human beings. The biochemical processes involved in the action of dextroamphetamine appear to be similar in spiders and in men (Peters et al., 1950), putting aside obvious differences in structural complexity. Web construction is a particularly narrow kind of behavior, regulated by sensory feedback but apparently unaffected by experience (Mayer, 1953; Petrusewiczowa, 1938; Reed et al., 1970). It does have the virtue of providing a rich and natural sample of behavior without encountering the difficulties which an experimenter confronts in choosing suitably representative and adequate behavioral indices for human behavior. Motivational problems such as the disinclination of the drugged human subject to tolerate experimental procedures are not encountered; the building of the web is a paramount requirement for the spider, making it highly likely that it will display any disabling effects of the drug. Unfortunately, it is not easy to identify a behavior which has a similar preferment in human beings. Given the difficulty in identifying independent maturational effects in the rather predictable, rigidly
70
REED, WITT, AND SCARBORO
patterned behavior of building webs, it is necessary to be cautious about drawing comparisons to changes in the complex behaviors of human adolescents and adults.
Notes
1 Requiring a web on each of six particular days, three of which inyolve drug effects, is stringent even for the high rate of building (approaching one web per day) which females of this species show at the period of life spanned in the experiments. (See Table 3.)
2 Loss of precision is more directly interpretable for spiral turns than it is for radii. In the process of attaching the spiral thread, clear probing movements of the first pair of legs can be observed; removing them disturbs the precision of the spiral (Reed, Witt, & Jones, 1965). For our radial precision measures, there is some inevitable apparent loss of precision because the spider does not fill the web space by a successive bisection of unfilled sectors. This spurious loss of precision would become greater as the number of radii fell below normal.
References
Aman, M. Q., and Singh, N. N. (1980). The usefulness of thioridazine for treating childhood disorders-fact or folklore? Amer. J. Ment. Def, 84:331-338.
Christiansen, A., Baum, R., and Witt, P. N. (1962). Changes in spider webs brought about by mescalin, psilocybin, and an increase in body weight. J. Pharm. Exp. Therap, 136:31-31.
Mayer; G. (1953). Untersuchungen ueber Herstellung und Struktur des Radnetzes von Aranea diadema und Zilla-x-notata mit besonderer Berücksichtigung des Unterschieds von Jugend- und Alters-netzen. Z. Tierpsychol. 9:337-362.
Koenig, M. (1951). Beitrage zur Kenntnis des Netzbaus orbiteier Spinnen. Z. Tierpsychol., 5:462-493. Peters, H. M., Witt, P. N., and Wolff, D. (1950). Die Beeinflussung des Netzbaues der Spinnen. Z. Vergl. Physiol., 32:29-44.
Petrusewiczowa, E. (1938). Beobachtungen über den Bau des Netzes der Spinnen. Trav. Inst. Biol.
Univ. Wilna, 9:1-25.
Reed, C. F. (1969). Cues in the web building process. Am. Zool., 9:211-221.
Reed, C. F., Witt, P. N., and Jones, R. L. (1965). The measuring function of the first legs of Araneus diadematus Cl. Behaviour, 25:98-119.
Reed, C. F., Witt, P. N., Scarboro, M. B., and Peakall, D. B. (1970). Experience and the orb web. Dev. Psychobiol, 3:251-265.
Witt, P. N. (1956). Die Wirkung von Substanzen auf den Netzhau der Spinne als Biologischer Test. Berlin: Springer Verlag.
Witt, P. N. (1961). Web building. In P. Gray (ed.), The Encyclopedia of the Biological Sciences. New York: Reinhold. Pp. 1072-1073.
Witt, P. N. (1971a). Drugs alter web-building of spiders. Behav. Sei, 76:98-113.
Witt, P. N. (1971b). Instructions for working with web-building spiders in the laboratory. BioScience, 21:23-25.
Witt, P. N., Reed, C. F., and Peakall, D. B. (1968). A Spider’s Web. Berlin: Springer Verlag,