Images Collection





View this article in Search Friendly Plain Text
NOTE: This plain text article interpretation has been digitally created by OCR software to estimate the article text, to help both users and search engines find relevant article content. To read the actual article text, view or download the PDF above.
Reprinted from
PHYSIOLOGY & BEHAVIOR
An Interndtiöfiijf
PERGAMON PRESS
NEW YORK • OXFORD • LONDON • PARIS
Physiology and Behavior. Vol, 3, pp. 119-124. Pergamon Press, 1968 Printed in Great Britain
Progressive Disturbance of Spider Web Geometry Caused by Two Sedative Drugs
CHARLES ^¿Í$ÉED AND PETER N. WIT||jf Department of Psychology, Temple University, Philadelphia, Pennsylvania
and
Division of Research, North Carolina Department of Mental Health, Raleigh, N.C.
(Received 19 Julm967)
Reed, C. F. and P. N. Witt. Progressive disturbance of spider web geometry caused by two sedative drugs. Physiol. Behav. 3 (1) 119-124, 1968.—The effects of diazepam and phenobarbital upon web-building show a time- and dose-dependency such that—in contrast to man—the action of phem|barbital is stronger than that of diazepam. Within the dose range here employed, the actions of both drugs are similar but have different rates of development. It is well possible, however, that if dose-levels were raised even higher, further differences would be found between the two substances. Both drugs produce significant curtailment of activity and ultimately evidence of imprecision of movement, presumably due to’ failure in sensory-motor coordination; this becomes apparent in radius construction for both drugs and in spiral placement for phenobarbital. Phenobarbital forces more radical economies of effort at lower dose-levels and earliSttimes than does diazepam. This curtailment of activity is not the potential thread mateiMfr at least, measure^, of gland
content under diazepam showed no slowing of glandular synthesis.
Spider web geometry Diazepam Phenobarbital
Die Wirkungen von Diazepam (Valium) und Phenobarbital (Luminal) auf den Netzbau von Spinnen zeigen Abhaengig-keit von Zeit und Dosis, wobei Phenobarbital (im Gegensatz zu Befunden am Menschen) die wirksamere Substanz ist. Innerhalb der hier verwendeten Dosierung sind die Wirkungen beider Substanzen aehnlich, aber entwickeln sich mit verschiedener Geschwindigkeit. Es ist aber auch gut moeglich, dass bei hoeherer Dosierung mehr Unterschiede in der Wirkung der beiden Substanzen auftauchen wuerden. Beide Substanzen bewirken eine signifikante Beeintraechtigung der Taetigkeit der Tiere, gefolgt von zunehmender Ungenauigkeit in den Bewegungen, wahrscheinlich als Folge des Versagens sensorisch-motorischer Koordinationen; dies zeigt sich im Radienbau nach beiden Substanzen und im Anlegen der Spirale nach Phenobarbital allein. Phenobarbital bewirkt auch eine weitgehende Einsparung des Kraftaufwandes bei niedrigerer Dosierung und zu fmeherer Zeit als Diazepam. Diese Beeintraechtigung der Aktivitaet ist nicht die Folge einer Veraenderung in der Menge bereitgehaltener Fadensubstanz; die Messung des Spinndrueseninhaltes nach Diazepam zeigt zumindest keine Verlangsamung der Fadensynthese Há|n Druesen.
One of the most striking characteristics of the orb-web of the garden spider, Araneus diadematus Cl., is its geometric complexity. It has been methodologically challenging to attempt to correlate structural components of this intricate product with the underlying behavioral and physiological systems. By manipulating the conditions of the spider—at first in rather gross ways such as by structural damage—some relations between patterns and behavior have been sketched out. It has been shown in previous work that the dimensions and geometric form of orb-webs are affected in specific manner by drugs which act upon the neural and muscular system of the spider (11). The array of changes seems characteristic for drugs or classes of drugs.
In the following investigation it was our intention to use drugs as tools to discover whether a common order of change and resistance to change could be detected in web-building behavior. If we interfere in the physiological functioning of spiders, we can study the behavioral consequences in the web-pattern. Drug effects are time related: they are known to appear and disappear gradually. Also, as the dose of the drug increases, it interferes increasingly with bodily function. Behavior, as measured in web patterns, should also change gradually. The two characteristics of drug action could therefore be used to study the sequence of web-changes as the physiological mechanism was more and more disturbed.
Web building of our spiders in the laboratory is fixed at a
*We thank Hoffmann-La Roche, Inc., Nutley, N. J. for giving us samples of diazepam, pure substance.
2The authors will be glad to provide detailed information or send a copy of the computer program on inquiry.
119
120
REED AND WITT
distinct hour in the early morning. By giving drugs at different intervals before this time, the effect of the drug can be followed over time. A slow acting drug, for example, given very briefly before web building, should have no effect on the immediately consequent web. If a drug reaches its peak effect in spiders after 36 hr, its application H days before the building of the test web would lead to maximal disturbances [9].
The spider’s pre-oral digestion permits application of drugs in watery solution by mouth. With a microsyringe the quantity of fluid which is attached to the spider’s mouth part can be exactly measured and adjusted to the body weight of the individual. Consequent observation can assure us that the drug was actually ingested. The concentration of the drug in the drop can be regulated so, that the dose required for that particular experiment is given to the animal under observation. This permits testing groups of spiders at different levels of drug concentrations.
It is apparent from these discussions that different experimental groups of spiders are needed to investigate the effect at several dose levels and several time intervals. Such an investigation demands a great number of data and calculations. This has only become possible after computer analysis has been introduced into the investigation of spider web patterns [5]. It is now possible to compare 27 parameters in hundreds of webs by grouping them according to experimental procedures, comparing the same webs of the same spiders before and after treatment.
Two drugs which possess sedative properties but which are chemically and therapeutically distinct were chosen for these experiments. Phenobarbital is one of the long-acting barbiturates with special anticonvulsant effects. Diazepam is classified by Jarvik [2] as a tranquilizer with muscle relaxant effects and a delayed onset of activity. The latter drug is given in man in l/100ths the dose of the former, indicating this drug to be the more potent substance.
Web-construction lends itself better than many other behavioral patterns to the assessment of severity of disturbance, because a web of some sort is nearly always built: it has been shown [7] that even after a high dose of caffeine the nearly paralyzed spider will construct some wild sort of thread pattern. It is surmised, therefore, and will be investigated in the following, that between such severe disturbances of web building and the normal web there can be a whole range of patterns with different degrees of alteration. In the end, we may learn something about what guides an animal through the elaborate process of orb construction, by gaining knowledge about the range of possible interferences in webbuilding.
METHODS
The webs of female cross-spiders {Arañeus diadematus) were photographed daily as described by Reed et al. [5]. The animals were 3-7 months old, and had been reared in the laboratory under controlled temperature.
No attempt was made to restrict the size of the animals used in the experiment, since the control webs were those built by each spider immediately previous to the drug-administration, and drug dose was adjusted to individual body weights. Body weight varies markedly even in spider-Jings of the same cocoon; therefore age is not a useful measure of animal size. The range of weights for these experiments was from 6-800 mg, with a median weight of 118 mg.
Some spiders were given drugs more than once, though never more than three times. At least two weeks intervened between drug-administrations to the same animal.
Under laboratory conditions, all spiders building a web on a given morning do so within a few minutes of each other. Drug administration was timed to precede the building-period by 12 and 24 hr. An interval of 24 hr must elapse before another web can be expected, hence the course of the drug effect in these experiments is measured conveniently at 12 hr intervals (up to 48 hr in one dose-level of diazepam).
Each animal was weighed and the volume of drug solution (or suspension) of known concentration adjusted, with the help of a Hamilton Microsyringe, so that an animal received the desired dose with an accuracy of better than 10 per cent. The spider was held while it drank in order to insure digestion of all of the fluid. However, it is possible that the animals later regurgitated some of the solution, particularly that of high concentrations with strong taste.
The two drugs, phenobarbital (Luminal) and diazepam (Valium), were dissolved (or suspended) as pure substances in enough sugar water to produce acceptance by the spider. Total volume of liquid per spider was about 10 p. Doses were 10, 100, 200 and 400 mg/kg of diazepam, and 10, 100 and 1000 mg/kg of phenobarbital.
For all drug levels for both drugs, the webs made by 20 animals prior to drug ingestion were compared statistically, measure by measure, with webs made by the same animals 12 and 36 hr after drug ingestion. The input to the computer were angles and distances measured on a web photograph; the computer calculated each measure and performed the statistics. Some of these experiments were replicated during building periods of two summers; where they are comparable, comparison of the two sets of data provides some information on the consistency of the findings. For one dose level of diazepam (100 mg/kg), control webs were also compared with webs built 24 and 48 hr after drug-ingestion. The extra dose levels were not replicated in the two summers.
Measures. Three general characteristics of each web were studied quantitatively: (a) the size of the several web-regions (distinguishing, however, between area and the number of threads found in a region) and including total thread length, (b) the shape of the main web, and (c) the precision of thread-placement. Each of these general characteristics is represented by several parameters, each differing in their relative contribution to the characteristic. For instance, total length of thread in the web, number of spiral turns, and number of radial threads are all measures of web size, considered from the point of view of expenditure of material. From a set of 100-200 selected data points measured on each web photograph, 27 parameters concerned with web-structure were extracted by a computer.’ Briefly, the measured points were the intersections of each spiral turn with-the radii at the four cardinal directions, and the location of the inner and outer limits of the spiral zone and the framing zone [5].
Each of the 27 parameters was tested for differences in a ¿-test of the 20 control and 20 experimental webs of each treatment. A chi-square analysis was also performed for the measures concerned with regularity of thread-placement, because of the non-parametric nature of the measure. We set significance at the one per cent level of confidence for all tests. In such a large number of tests, spurious significance effects are to be expected with a frequency set by the significance level; the replicated summers assisted in evaluating such circumstances.
PROGRESSIVE DISTURBANCE OF ¡SPIDER WEB GEOMETRY
Apart from these measures concerned with the geometry of the web, it was also necessary to evaluate the spider’s use of the material available in the ampullate glands; these glands produce all the scaffolding for the web [3], A sample of webs which appeared to be smaller or sparser (as a result of the drug) was digested; nitrogen content was determined in the digest in order to establish the amount of material expended in construction (Method, see 1).
RESULTS
Diazepam
Thread Expenditure
Curtailment of expenditure of thread occurred for all dose-levels of diazepam used in the experiment.
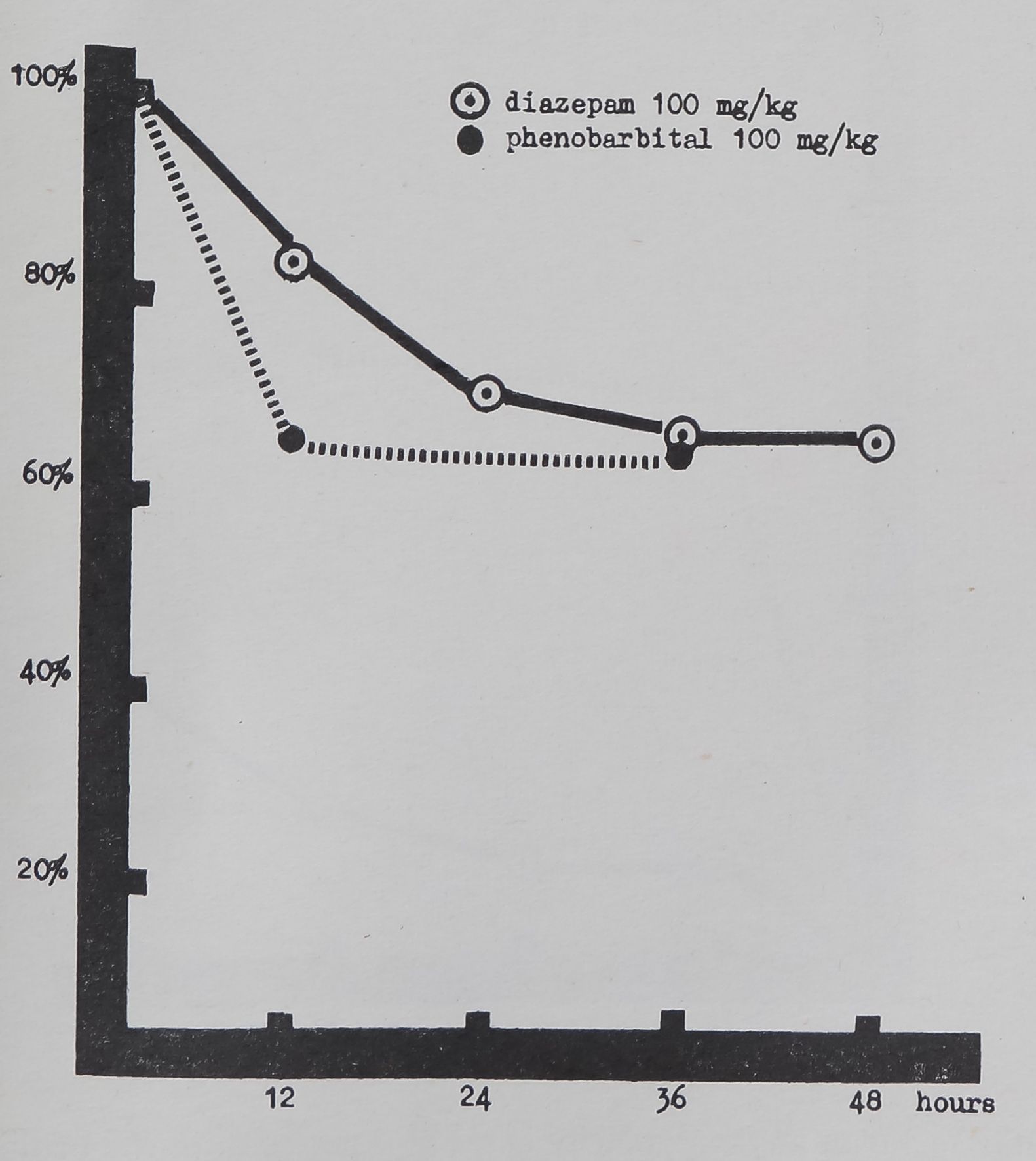
Thread length. There is significant reduction in the total length of the lines composing the web (Table 1 and Figs. 1 and 4B). The effect appears at all dose-levels and in early and late webs; it lasts at least through 36 hr (the 100 mg— 48 hr sample showed below 5 per cent significance.J ‘
Web area and thread elements. The curtailment of thread in the experimental webs appears to be a matter of reduction of the number of thread elements rather than a “shrinkage” of the space occupied by the web. The threads of the main web, those forming the viscid spiral or catching zone, are consistently fewer in number on the drug webs; on the other hand the area covered by the spiral threads shows only occasional reduction in magnitude (Table 1 and Figs. 4A and B).

FIG. 1. Mean thread length in 40 control webs was assumed as 100 per cent. Decrease in length is plotted in per cent of control on the ordinate; abscissa represents time between drug application and web-building. Both 12-hr, the 24-hr and both 36-hr values are significantly below control values.
Note the relatively slow onset of action of diazepam in comparison to phenobarbital and the long duration.
121

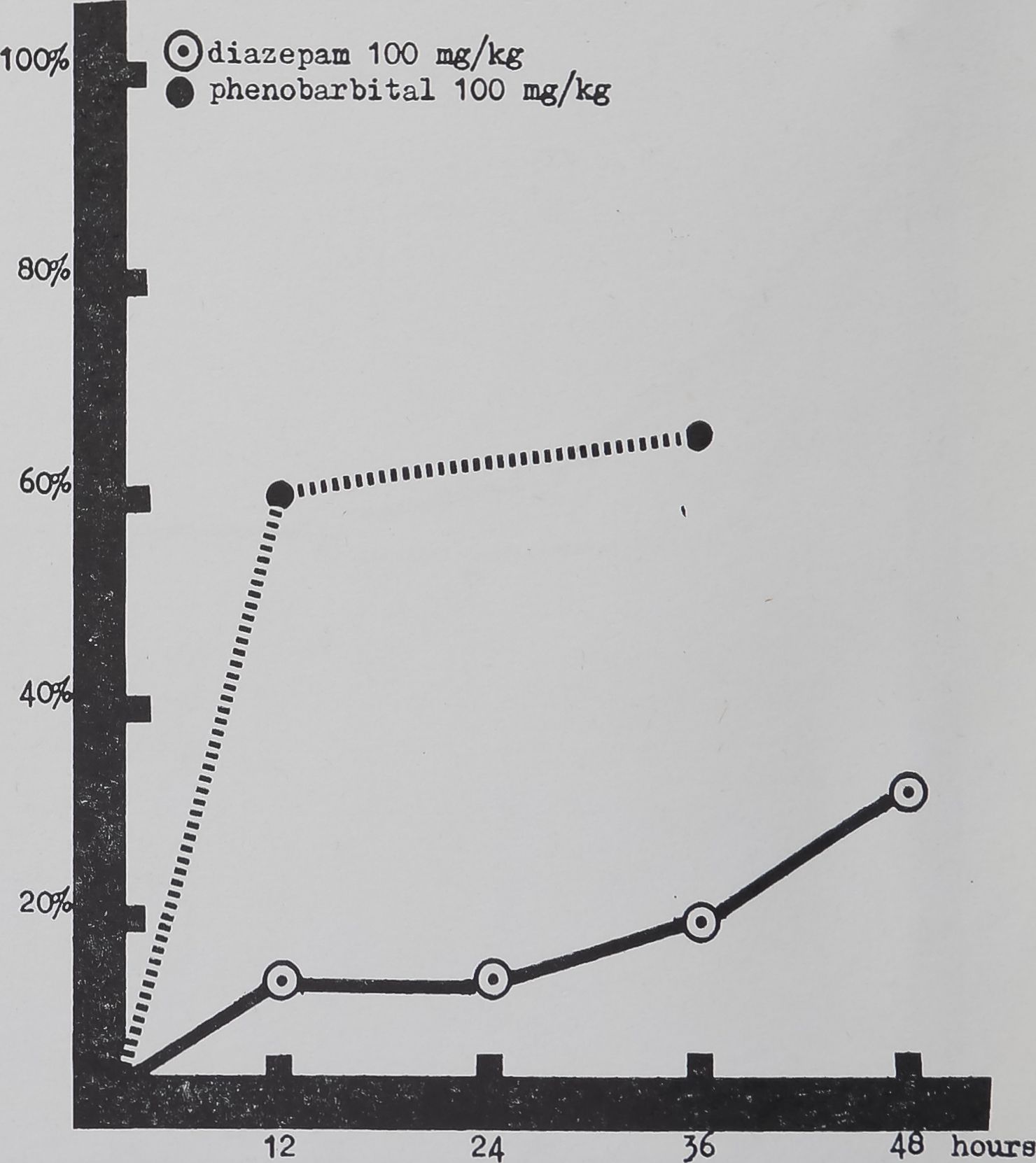
FIG. 2. Per cent change in mean number of radii per web in 40 webs of Araneus diadematus Cl. is plotted on the ordinate against time of drug action on the abscissa. The 24 and both 36 hr values are significantly different from controls.
Note the distinct but statistically not significant decrease in mean
number of radii in webs built 12 hr after drug application.
“Radial” threads,! ie. those lines which run from the hub in a radial pattern to the web periphery, decline significantly in number for diazepam doses above 10 mg/kg (Fig. 2). The effect never appears in the 12 hr webs, however, and is absent in the samples whieh were taken at 48 hr for the 100 mg dose level and at 60 hr for the 400 mg dose level. There appears to be an order in which the radial and spiral threads are affected by the drug: a slower “penetration” into the mechanisms underlying placement of radial lines than those underlying spiral-building.
Thread material. Are the foregoing changes in number of threads and in total thread length a consequence of drug-action on the source of silk production ? It has been observed that webs which differ in length and number of thread elements can contain the same amount of thread material. Christiansen et al. [1] found evidence of relatively thick threads being spun by spiders which had weights attached to the abdomen. Atropine affects the amount of material present in the webs by its action on protein-synthesis in the thread glands [4, 8].
To answer the question in the present case, we measured total nitrogen in the web digests (as an index of quantity of thread material) in control webs and in webs built 12, 36 and 60 hr after application of 100 mg/kg diazepam. The nitrogen per web decreased from a control level of 35 units per web (N =Hl7) to levels of 21, 20 and 22 respectively for the three postdrug periods. Hence the webs built under diazepam contained less material than did control webs. Thread length decreased for the same periods from a mean of 14 per web to 11, 12 and 11 m, respectively. A rough calculation showed that there was, if anything, a slight decrease in thread thickness.
The question remains whether less material was available at web-building time or the glands were not emptied as usual. Peakall [4] has shown that drugs can decrease protein-synthesis at the cellular level in isolated ampullate glands; the rate of synthesis was deduced from the amount of material which could be obtained by emptying of the glands by pulling thread. In order to determine whether diazepam had a direct effect on synthesis, the amount of silk which could be pulled from the ampullate glands on control days (N = 13 spiders) was compared with the amount of silk which could be pulled one and two days after application of
122
REED AND WITT
TABLE 1
Changes in Architecture of Webs of Araneus diadematm following administration of ‘‘Diazepam”, listed by Dose-Level and Elapsed Time. Presence of an Entry indicates Departure from Control Values reaching at least the 0.01 Level of Significance
(t AND CHI-SQUARE TESTS). FOR HöH COMPARISON, EXPERIMENTAL AND CONTROL WEBS OF AT LEAST 20 ANIMALS WERE USED.
| Size measures | Regularity measures | ||||||
| Dosage
(mg/kg) |
Hr | No. radii | No.
spirals |
Areas Frame Spiral | Thread
Length |
Area
Ratios |
Angle Spiral Mesh size |
10 12
| 36 | 9SBf| | Fewer | 9E£^ | — | Shorter | Irreg | ■ | |||
| 100 | 12 | — | Fewer | — | ?9BB | Shorter | — | — | ||
| 24 | Fewer | Fewer | — | — | Shorter | F/S*> | — | — | — | |
| 36 | Fewer | Fewer | —j | Smaller | Shorter | — | — | — | ||
| 48 | 99| | — | — | H^9 | Larger SEf | |||||
| 200 | 12 | — | Fewer | — | ^^9 | Irreg. | — | ■BH | ||
| ‘. 36 | Fewer | ; Fewer | Shorter | # “V | Irreg. | — | Larger SEf | |||
| 400 | 12 | 99| | Fewer | Smaller/,1 | Shorter | F/S*> | Irreg. | 9 | 99fl | |
| 36 | Fewer | Fewer | — | Shorter | 91 |
* Frame over spiral, f Standard error.
Not listed in this table are additional parameters wffim failed H change at any treatment level: shape of the main web, number of “oversized” angles, area of Central zone dB. hub and “free” zone) and mesh size;(two measures).
The enfries under angle regularity represent llpffeal measures: size of median angle, standard deviation of central angles, and a measure based upon size-differences between adjacent angles*
I^Bie number of spiral turns varies by quadrant in the normal web; entry is made in this table if significant change f¡|number occurs for at least one BBmtjÉ.9
100 mg/kg diazepam. The mean nitrogen value for pulled thread was 26 units per spider; for the first and second days after the drug it was 25 and 27, respectively. We interpret this result to indicate that the drug did not affect protein-synthesis, and that reduction in expenditure was not for want of thread material.
Precision of placement. The foregoing results are all interpretable as reflecting reducti om| the activity level of energy output of the animal. The reduction is selective, in the sense that spiral number is diminished, but not necessarily the area of space covered by the threads. The area of the zone is preserved, but more sparsely subdivided by thread.
The normal web, however, I# characterized by a certain degree of regularity in spacing of radial angl spiral threads; they bound approximately equal angles and shapes, especially if they are adjacent. Statistical test for departure from that regularity produces significant and near significant angle irregularities for several experiments with diazepam, but clear irregularity only at 200 mg or more per kilogram (Fig. 3). ¡Spiral regularity, on the other hand is at normal values (Table 1 and Fig. 4B).
Diazepam did not produce change in the overall shape of the web.
PHENOBARBITAL
The foregoing description permits us to briefly enumerate the effects of phenobarbital.
Thread Expenditure
The phenobarbital webs are significantly reduced, at all dose levels, in thread length, number of spirals, number of

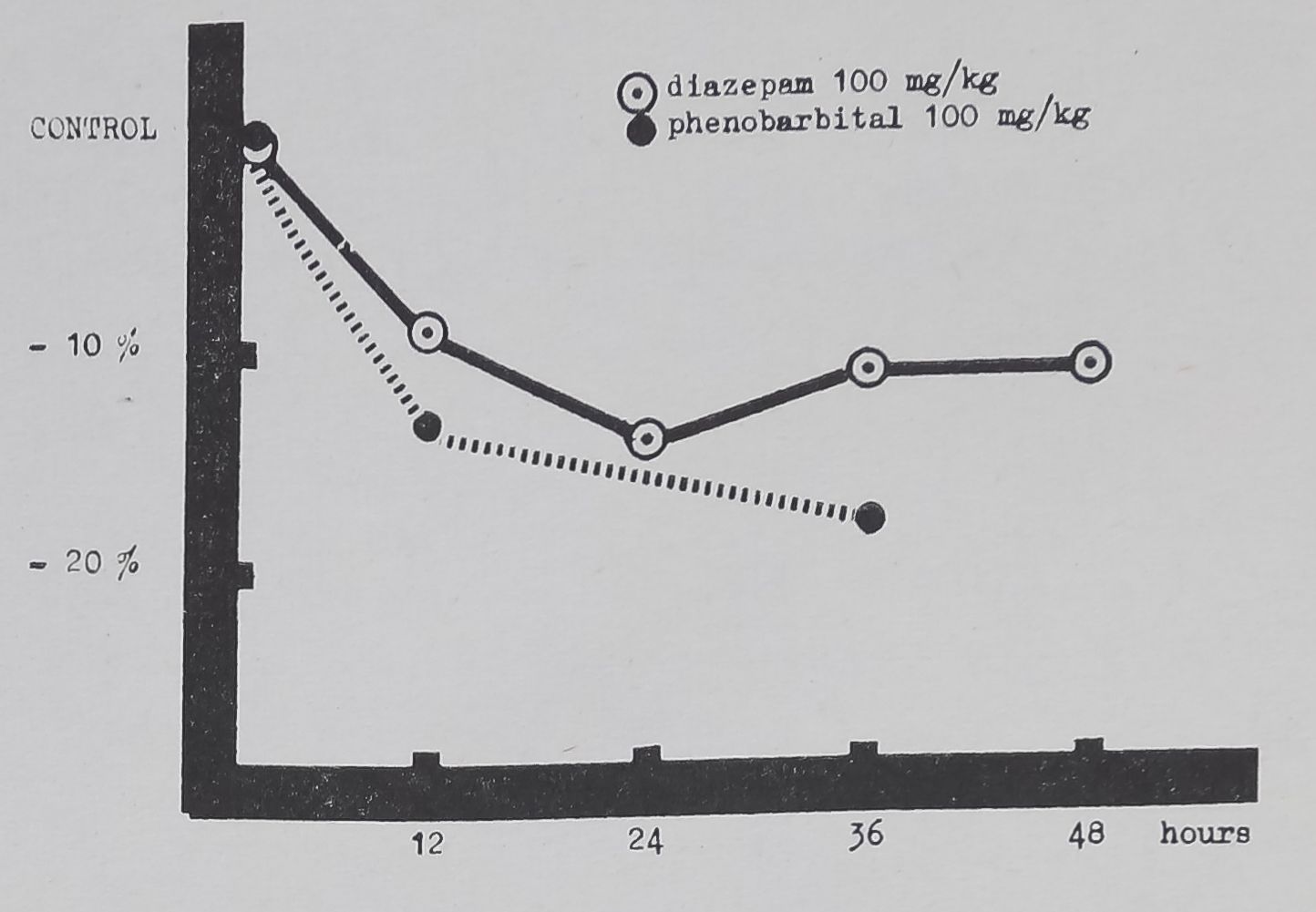
FIG. 3. Mean angle regularity is set at 0 per cent on the ordinate, an increase representing more irregular central angles; abscissa as in Figs. 1 and 2. Only the 12 and 36 hr phenobarbital values are significantly above control means.
Observe the steady small increase in irregularity after diazepam which does not reach statistically significant values with this dose during the period of our observations, in contrast to the rapid, significant increase after phenobarbital.
FIG. 4B. The same spider built this web 4Jp^later, 36 hr after it FIG. 4A. Control web of adult female Araneus diadematus Cl. ■ mBíÉ 100 mg/kg diazepam.
The straight white lines in the upper right corner indicate the Observe the decrease in number of radii and spiral turns and original distance || 20 mm and direction of gravh^|fe|i length of thread .(jtqtal path) as shown in Ta||l»M|
(,facing page 122 )


FIG. 5A. Control web B adult female Araneus diadematus Cl. The irregularity of spiral placement in the upper left corner is the result of the spider’s being forced to run out of the web prior to photography.
FIG. 5B. This web was built by the same spider as 5A, 24 hr later, 12?$fr after it had received 100 mg/kg phenobarbital. Accidental disturbance of spiral left as in 5A.
Observe the characteristic decrease in thread number and length and some irregular central angles.

FIG. 5C. This web built by the same spider 48 hr after web in 5A and 36 hr after 100 mg/kg phenobarbital, contains somewhat more severe changes in the pattern after the longer time. Compare also Figs. 1, 2 and 3.
It should be noted that even these statistically highly significant alterations in the web pattern are hardly apparent to the casual observer of this still beautifully patterned orb-web.


PROGRESSIVE DISTURBANCE OF SPIDER WEB GEOMETRY
123
TABLE 2
Changes in Webs of Áraneus diadematus following Administration of “Phenobarbital” at the Doses listed and for two Building Periods. Presence of an Entry indicates Departure from Control Values reaching at least the 0.01 level of Significance (t and
CHI-SQUARE TESTS). FOR EACH COMPARISON, EXPERIMENTAL AND CONTROL WEBS OF AT LEAST 20 ANIMALS WERE USED. (Parameters are described further in legend of Table 1.)
Size Measures Regularity Measures
| Dosage
(mg/kg) |
Hr | No.
Radii |
No.
Spirals |
Areas Frame Spiral | Thread
Length |
Area
Ratios |
Angle | Spiral | Mesh size | |
| 10 | 12 | Fewer | Fewer | Smaller | Smaller | Shorter | ,__ | Irreg. | ||
| 36 | Fewer | Fewer | 1 Hj I | Smaller | Shorter | C/S*> | Irreg. | Irreg. | ||
| 100 | 12 | Fewer | Fewer | 1 | Smaller | Shorter | — | Irreg. | — | Irreg. |
| 36 | Fewer | Fewer | — | Smaller | Shorter | — | Irreg. | — | Irreg. | |
| 1000 | 12 | ‘ — | Fewer | Smaller | — | — | — | Irreg. | ||
| 36 | Fewer | Fewer | Smaller | Shorter | Irreg. | Irreg. | ||||
♦Center over spiral.
radial threads and spiral area (Table 2 and Figs. 1, 2, 5A, B, Q. These effects last through the 36 hr webs.
Precision of Placement
Irregularity of central angles appeared in the lowest experimental doses and the earliest constructions, it is still present in the 36 hr webs (Fig. 3). Spiral irregularity appeared only in the webs of the high-dose level, 1000 mg/kg.
Shape of the phenobarbital webs did not depart from control values.
DISCUSSION
Both diazepam and phenobarbital produce a significant diminution of the activity level of the spider (as evidenced by reduced thread length and number of spiral turns) without affecting the amount of material available for the web. The decline in activity is more radical for phenobarbital than for diazepam: disturbances which appear early and at low doses for phenobarbital are present only in the late and high dose-levels of diazepam. This is in contrast to man, where diazepam appears to be the more potent drug. We have no evidence at this time which would favor one particular explanation of this finding.
The size of the main web (the spiral zone) and the number of radii are diminished in the 12 hr webs for phenobarbital at all doses; spiral zone is not so clearly affected for diazepam, and radial number is never reduced in the 12 hr webs. Angle irregularity occurs clearly in the phenobarbital webs, but rather less consistently in the diazepam webs.
Spiral irregularity, which is present for the high doses of phenobarbital does not appear in the diazepam records; whether it may do so at higher doses of diazepam cannot be answered from these experiments.
Figures 1, 2 and 3 compare the reactive development of effects for the two drugs.
We conclude that phenobarbital produces a decline in level of activity and in sensory-motor control the latter being only intimated at high levels of diazepam, using the terms “high” and “low” to refer to the dose-levels employed in these experiments. Neither drug affected the shape of the web, a disorder which can be observed for other drugs, and which seems to represent marked sensory-motor disturbance.
An order of vulnerability to disturbance of web-building is suggested by the time and dose-dependent results. The changes in web dimensions and pattern, reflecting progressive handicap to the functioning of the spider, may reflect the sequence of events for diazepam or phenobarbital alone. On the other hand, the accumulation of disturbances may be distinctive for sedative drugs generally. The question is one for subsequent experimentation; the order indicated by the present work falls into three stages:
1. Curtailment of activity. The available silk is not emptied from the gland, as appears to be the case in normal building [3]. A smaller than normal amount of material is utilized because the spider traverses less space than usual (thread emission invariably accompanies movement). The total path, as measured by length of thread is reduced (a) without loss of precision of placement of threads (adjacent central angles and adjacent spaces between spiral turns remain approximately equal) and (b) without reduction in the area of the spiral zone (threads are sparsely distributed, but the limits of the zone remain at control dimensions).
The threads composing the spiral are diminished in number in at least one quadrant before the radial threads are affected.
2. Reduction of area of the spiral zone. The dimensions of the spiral zone are contracted. Precision of placement of threads is preserved, but because of the smaller area of the essential sticky trapping region, the web is less effective as a snare than is the normal web.
3. Loss of precision of placement. There are two substages; (a) the central angles of the web become more variable, less regular than normal, (b) spaces between spiral turns become less regular than normal. The precedence of angle irregularity over spiral irregularity which occurs for phenobarbital is consistent with the comparative complexity of the building processes involved in these structures. In the laying of the spiral, the spider is apparently guided by touching structures already made; construction of the radial threads, however, occurs as one of the first events of web-building, when there are in a sense many more “degrees of freedom”. Complex testing and guidance functions are operative at that time; their complexity is consistent with greater vulnerability to disturbance.
124
REED AND WITT
In the case of phenobarbital, then, angle irregularity precedes spiral irregularity. It is possible that the radial threads could be so irregularly arrayed that it would be difficult if not impossible to lay a regular spiral upon them. That degree of disturbance did not occur for the dose-levels used in this experiment. For diazepam, the higher standard error of mesh size may indicate a slight disturbance in spiral placement. However, at the dose levels employed, diazepam causes no detectable change in the more direct measure of spiral regularity (Table 1). This raises the pharmacologically interesting question whether spiral regularity is sensitive to one and not to the other sedative drug, or whether the maximum diazepam dose employed was insufficient.
The order of substages may be specific to these two substances. For example, a different order is observed after amphetamine where disturbance in spiral occurs prior to disturbance in central angles [10]. There are also form-changes which have been observed in other types of experimental treatment but which did not appear under diazepam and phenobarbital, e.g. disturbance of position of the hub and of the characteristic oval shape of the web. This schema is therefore provisional and intended to be heuristic to the observation of change sequence in other drug-treatments.
Limitations and Advantages of Method
Not only the choice of a relatively unusual experimental animal and its particular behavioral repertoire impose restriction on the interpretation of our results: the selection and statistical processing of measures of web pattern are based upon arbitrary decisions which must be rationalized.
The number of statistical tests performed (27 tests per dose-level and time-period) enhances the probability of obtaining significant difference. To compensate for this circumstance, we have chosen to make errors of Type II, i.e., to miss real differences, by setting a somewhat stringent level of significance. We have also examined the results of the two sets of summer experiments for consistency of significant differences for all measures. Not all experiments were repeated in both summers, and modifications were made in the second series on the basis of experience in the first. It
was found for instance that the first series extended too far into late summer when the animals were beginning to show seasonal decline in frequency of web-building. While comparison is limited for these reasons, there are 205 pairs of ¿-tests in which it is justifiable; 21 of these reached the 0.01 level of significance in one summer but not in the other. The direction of change was consistent however; pooling the data for the two summers produced 0.01 level significance in 20 of the pairs. The experimental differences found for spiral number, thread length, angle regularity, radial number and spiral zone are particularly consistent.
A few comments may be made in defense of choice of experimental animal. In behavioral investigation, it is often difficult to distinguish between an incapacity to perform and a disinclination to do so. In order to demonstrate motor disfunction for instance, an experimenter must contrive procedures to insure that his subject is adequately motivated to act at all. A drug may produce loss of motivation through malaise or other states which interfere with performance in a very general sense. The more elaborate or precise the behavior, the more subtly do motivational states intervene.
Web-building requires refined execution of motor-sequences —all capable of large and small disturbances—but it occurs even under conditions in which the animal is severely handicapped. It is not only that motivation is rarely ascribed to spiders, but that the apparently strong necessity to build— however this necessity comes about—provides the experimenter with action more likely to be modified than terminated. Both diazepam and phenobarbital do produce some decline in web-building frequency: as dose-level is increased, the number of animals building on both of two consecutive building-periods declines. This trend does not reach statistical significance, although it is doubtless possible to produce general anesthesia with drugs [6]. This possibility, however, does not affect the point that if any movement at all is possible for the orb-weaving spider it is likely to attempt to build its intricate web at the appropriate time of day.
This work was in part supported by a grant from the National Science Foundation. The assistance of Mrs. Marguerite Agi is gratefully acknowledged.
REFERENCES
1. Christiansen, A., R. Baum and P. N. Witt. Changes in spider webs brought about by mescaline, psilocybin and an increase in body weigh^/. Pharmac. exp. Ther. 136: 31-37,1962.
2. Goodman, L. S. and A. Gilman. The pharmacological basis of therapeutics, 3rd Ed. New York: MacMillan Co., 1965,107-187.
3. Peakall, D. B. Composition, function and glandular origin of the silk fibroins of the spider Araneus diadematus Cl. J. exp. Zool. 156: 345-352, 1964.
4. Peakall, D. B. Regulation of the synthesis of silk fibroins of spiders at the glandular level. Comp. Biochem. Physiol. 15: 509-515, 1965.
5. Reed, C. F., P. N. Witt and R. L. Jones. The measuring function of the first legs of Araneus diadematus Cl. Behaviour 24: 98-119, 1965.
6. Schwarz, R. Versuche ueber die Beeinflussung des Netzbaues der Spinne Zilla-x-notata CI. durch Stickoxydul und Aether. Archs. int. Pharmacodyn. Ther. 104: 339-364, 1956.
7. Witt, P. N, Verschiedene Wirkung von Pervitin und Coffein auf den Netzbau der Spinne. Helv. physiol, pharmac. 7: C65-C66, 1949.
8. Witt, P. N. Effects of atropine on spiders’ web building behavior and thread production. Fedn. Proc. 21: 1962.
9. Witt, Peter N. Effects of Physostigmine on spiders’ web building behavior and thread production. Pharmacologist, Fall, 1963.
10. Witt, P. N., L. Brettschneider and A Boris. Sensitivity to d-amphetamine in spiders after iproniazid and imipramine. J. Pharmac. exp. Thér. 132: 183-192, 1961,
11. Witt, P. N. and C. F. Reed. Spider-web building. Science 149: 1190-1197, 1965.