Images Collection











Read OCR Digitized Article Text
NOTE: This plain text article interpretation has been digitally created by OCR software to estimate the article text, to help both users and search engines find relevant article content. To read the actual article text, view or download the PDF above.
Behavior Genetics, Vol 7, No, 3, 1977
Quantitative Analysis of Orb Web Patterns in Four Species of Spiders
Peter Risch1,2
Received 18 July 1975—Final 18 May 1976
The orb webs of the four species Araneus pima (species nova), Araneus diadematus (Clerck)$ Argiope aurantia (Lucas), and Neoscona domicile orurrr (Hentz) built under controlled laboratory conditions were compared and evaluated quantitatively at two different points in the life cycle in regard to size, regularity, shape, and fine structure. Orb webs of two species from one genus can be distinguished, but are more alike than webs from different genera. Mature spiders built highly specific webs, a fact which may play a role in species isolation during courtship. In adults, web measures correlated in size with the spiders’ taxon; young spiders’ web size measurements corresponded rather to their body weight or leg length. A web’s regularity seemed to be closely related to age. Adult and young spiders built oval-shaped webs with eccentric hubs. Mesh width was correlated with leg length at both age levels. Web data support the likelihood of a monophyletic evolution of the four species.
KEY WORDS: taxonomy; invertebrate behavior; maturation; web patterns; spider webbuilding.
INTRODUCTION
The spider’s web, as compared to the results of other animals’ patterned movements, presents a most favorable record for quantitative behavior
This work was supported by Grant GB-25274 from the National Science Foundation awarded to Dr. P. N. Witt, and was carried out in the laboratories of the Division of Research, North Carolina Department of Mental Health.
1 Research Division, North Carolina Department of Mental Health, Raleigh, North Carolina.
2 Present address: Zweierstrasse 186, Zurich 80Ô3, Switzerland.
199
© 1977 Plenum Publishing Corp., 227 West 17th Street, New York, N.Y. 10011. To promote freer access to published material in the spirit of the 1976 Copyright Law, Plenum sells reprint articles from all its journals. This availability underlines the fact that no part of this publication may be reproduced, stored in a retrieval system, or transmitted, in any form or by any means, electronic, mechanical, photocopying, microfilming, recording, or otherwise, without written permission of the publisher. Shipment is prompt; rate per article is $7.50.
200
Risch
analysis. Comparison of similarities in web details of various spiders can provide data for ontogeny and phylogeny of behavior.
Many spiders build daily orb webs with a characteristic geometric pattern. This geometric arrangement, an expression of movements in time and space, is a relatively permanent record of a significant part of the behavior of the thread-laying animal; it contains an enormous amount of information, which can be defined, measured, and evaluated. Certain web features are correlated with age, size, weight, sex, and species (Witt et 1972). It is surprising that these measurable records were seldom used for quantitative spider classification.
Spider systematics has been pursued from a morphological point of view. Many different body characteristics have been identified and serve as determinants of species, families, and genera, which in turn determine phylogenetic relationships. T)o we find the same classification in using behavior data resulting from web measurements as we do with morphological criteria? This will be a basic question in this study.
I compared the web-building patterns from two species of one genus and one species each of two other genera at two different times of life, i.e., directly after the last molt and at the midway point between hatching and the last molt. The spiders were compared at two different periods in life to identify changes in web patterns during ontogeny as well as to determine if differences among genera, species, and broods (the term “brood” is used in this article to define spiders which hatched from the same egg sac) are more apparent later in ontogeny. These comparisons permitted conclusions as to the time in the spider’s life at which the orb web is a more distinct expression of the genetic background. Animals of comparable development were selected for web comparisons, rather than animals of equal weight or similar leg length, because one may assume that our four species at similar stages of development also built their webs for similar functions. The function of the web in prey catching, in enlarging the perceptual range, and in providing a warning system, runway, and platform for courtship should be optimal at maturity. Perhaps males are attracted to resonant properties of webs of their own species. If so, this may work for species isolation. One would then find highest species specificity of webs at maturity. If webs play no significant role in species isolation at mating time, mature web and young web could be equally unspecific. I also deal with the question: How far can a morphological characteristic (i.e., leg length) of a species determine behavior (i.e., leg movements, which in turn determine density of fine structure)?
The pattern of a spider’s web is the result of a behavior program in the nervous system rather than being dependent on experience (Reed et al., 1970); this program, in turn, is an expression of an inherited genetic code. Is
Quantitative Analysis of Orb Web Patterns in Four Species of Spiders
201
the code for web building a result of the gene pool pressure of the ancestral background or is it rather an individual adaptive response to the particular environment? In the first case, one could suspect that intrageneric webs show less differences than intergeneric webs, even when species from the same genus occupy different habitats with distinct prey behavior and size. In the second case, we may assume that species which occupy similar habitats show some comparable web characteristics even when they belong to different genera. The present study will be concerned with this problem, too.
Interpretations of phylogenetic relationships between different web types and their evolution have been made for many web builders by several authors on a speculative basis (Kaston, 1964; Kullman, 1958; Peters, 1939; Szlep, 1966; Wiehle, 1929). It will be done quantitatively on a few selected species in this study with regard to web ontogeny, phylogeny, and adaptation.
All experiments were done under similar laboratory conditions. Consequently, differences in web patterns found among broods, species, and genera are due to genetic variations rather than to contrast in the environment.
MATERIALS AND METHODS
The webs considered in this study were build by four species of spiders, belonging to three genera (Table I and Fig. 1). Araneuspima and Neoscona domiciliorum were identified by Dr. W. J. Gertsch (Portal, Arizona 85632), Araneus diadematus and Argiope aurantia by Dr. H. W. Levi (Harvard University, Cambridge, Massachusetts 02138). The total number of broods was seven. From the time of hatching to the end of the experiment, the
Table I. Spiders Whose Webs Were Used for Analysis, Their Place of Origin, Their
Hatching Date®
| Genus and species | Brood | Egg sac origin | Hatch date |
| Araneus pima | Aspl | Portal, Arizona | 14 Jan. 1974 |
| Araneus diadematus | Asdl | Canastota, New York | 25 Feb. 1970 |
| Asd2 | Canastota, New York | 22 Apr. 1970 | |
| Argiope aurantia | Aeal | Raleigh, North Carolina | 8 Dec. 1969 |
| Aea2 | Raleigh, North Carolina | 8 Mar. 1970 | |
| Neoscona domiciliorum | Nadi | Knightdale, North Carolina | 1 Nov. 1971 |
| Nad2 | Knightdale, North Carolina | 18 Jan. 1972 |
The offspring of a single cocoon are called a “brood.”
202
4
Risch
a
I
Fig. 1A-D. Four mature female spiders from the different species whose webs were compared. A: Araneus pima. Age 292 days. Body weight 500 mg. Total leg length 21 mm. Sternum, carapace, and legs brown. Abdomen gray-brown. Note the two large humps. For Araneus pima’s distribution, habits, and detailed description, see Levi (1971). B: Araneus diadematus. Age 251 days. Body weight 405.mg. Total leg length 20 mm. Sternum dark brown, carapace and abdomen yellow-brown. The legs banded: yellow-brown with dark brown. Note the white marks at the anteriorly dorsal end of the abdomen in the form of a cross, which identifies the species. For distribution, habits, and description, see Levi (1971) or Grasshoff (1968).


Quantitative Analysis of Orb Web Patterns in Four Species of Spiders
203
Fig. h Continued. C: Argiope aurantia. Age 286 days. Body weight 303 mg. Total leg length 30 mm. A median longitudinal yellow-white band divides the black sternum. The carapace looks yellow-white and shows some brown markings. The dorsum of the abdomen is marked black and yellow. Note the silver-white hairs on the carapace and the dark brown banded legs. Compare the morphological features, legs, carapace, and abdomen with those of the other three species. For distribution, habits, description, see Levi (1968). D: Neoscona domiciliorum. Age 260 days. Body weight 201 mg. Total leg length 15 mm. The abdomen shows a characteristic pattern with yellow or white colors of the anterior dorsal surface and transverse bars on both sides of the posterior part. Note the distinctive dorsal maculation. For distribution, habits, and description, see Berman and Levi (1971).


204
Risch
spiders were kept in a room with programmed lighting: 8 hr dark alternating with 16 hr light. High humidity was obtained through a room humidifier, which was in operation at all times. The temperature was always 5°C lower during the dark hours than during light and increased each morning (average night temperatures 15-18°C, day temperatures 20-25°C).
The origin of the egg sacs and their hatching dates are shown in Table I. The cocoons were brought to the laboratory, and as soon as the spiders hatched they were placed in a rearing box, where they built a communal web. The spiders were fed a constant supply of gnats until they started to build individual orb webs. Then they were removed to aluminum and glass cages, 50 by 50 by 10 cm (Witt, 1971). Only one spider was kept in each cage, so that the spiders were identifiable throughout life. From then on, they were fed twice a week with one housefly and watered 5 days per week by spraying water into the cage.
About one web per spider was photographed each week. For photography, the glass doors of the spider’s cage had to be removed without destroying the web. Then the web was placed in front of a black box, the opening surrounded by a square of fluorescent light tubes. Between subject and camera, a mask (movable screen) was placed in such a way that only light reflected by threads could reach the camera. In order to get an oriented and scaled reproduction from the negative, a scale of two threads, 20 mm apart, and a metal bar were placed near each web (see Figs. 2 and 3). The web was photographed with 35-mm Kodak Plus-X film (ASA 125), 0.5-1 sec exposure time (depending on web-to-camera distance) and a diaphragm opening of f/3.5 to f/5.6. A detailed description of web photography is found in Peters (1970) and Witt (1971).
By means of an enlarger, the negatives of web photographs were projected to original size and position. Approximately 200 measurements were obtained for each web. These selected data were punched on cards and processed by computer. For each web, 27 variables specifying dimensions in size, regularity, shape, and fine structure were calculated. Detailed information about calibration of photographs and the deriving of web measures and their function is found in Witt et al. (1968) and Witt (1971).
In this study, four dimensions of size (Tables IV and V), four of regularity (Tables VI and VII), three of shape (Tables VIII and IX), and one of fine structure (Tables VIII and IX) were compared among three genera, four species, and seven broods (Table I). The behavioral meanings of the four different dimensions are explained in the results. Table II shows how many spiders were studied from each brood, as well as the number of webs for each sample. In order to obtain comparable numbers of webs for each species, only 40% of the many available Araneus pima webs were randomly selected, while 90% of the available webs of the other broods were chosen.
Quantitative Analysis of Orb Web Patterns in Four Species of Spiders
205

Fig. 2A-D. Four webs built by immature female spiders. The two vertical white lines in the upper left corner were originally spaced 20 mm apart; they indicate also the vertical direction in the original web. Compare the four webs with the data in the Tables II, IV, VI, and VIII. Note differences in scale. A: Web of Araneus pima. Spider’s description at time of web-building: age 100 days, body weight 35 mg, leg length 8 mm. Web measures: spiral area 29,200 mm2, center area 2350 mm2, thread length 12,211 mm, angle regularity 1.8°, mesh width 34.3 mm2. Note the high angle regularity and the circular web form (width/length) as well as the irregular tangle of threads at the upper right corner of the frame. These thread structures, which are typical for most immature and all mature Araneus pima webs, are not taken into consideration in our measurements. B: Web built by Araneus diadematus. Spider’s description: age 45 days, body weight 38 mg, leg length 8 mm. Web measure: spiral area 31,938 mm2, center area 3314 mm2, thread length 15,991 mm, angle regularity 3.1°, mesh width 43.5 mm2. Note the relatively big spiral and center areas, as well as the elliptical web form. C: Argiope aurantia built this web. Spider’s description: age 120 days, body weight 25 mg, leg length 12 mm. Web measures: spiral area 20,674 mm2, center area 2449 mm2, thread length 8227 mm, angle regularity 3.0°, mesh width 58.9 mm2. Note the species-specific silver-white “hatched band” or stabilimentum. This structure is not considered in this study. Compare leg length and mesh width. D: This web was built by Neoscona domiciliorum. Spider’s description: age 75 days, body weight 20 mg, leg length 8 mm. Web measures: spiral area 31,388 mm2, center area 1535 mm2, thread length 15,419 mm, angle regularity 3.1°, mesh width 38.1 mm2. Note the small center area in relation to the big spiral area.


Quantitative Analysis of Orb Web Patterns in Four Species of Spiders
207

Fig. 2. Continued.
The differences among genera, species, and broods were tested by oneway ANOVA. Comparisons between juvenile and mature spiders were done by t test for paired samples (Table X). Differences between pairs of means were tested with the LSD test (least significant differences); capital letters were used to visualize the differences (see Table II).
The mature spiders from the four species used in this study showed different morphological characteristics (Fig. 1). The same discrepancies in morphology, body weight, and dimensions in size, such as leg length, could also be seen in the young spiders (Table II). Under these circumstances, it was impossible to select homogeneous spider material in each regard. For reasons which are described in the introduction, I decided in this experiment to have four species with comparable degrees of development. At two points in the life cycle, webs from spiders with similar maturity were collected and compared. The mature spider webs, three for each spider, were built in the 3
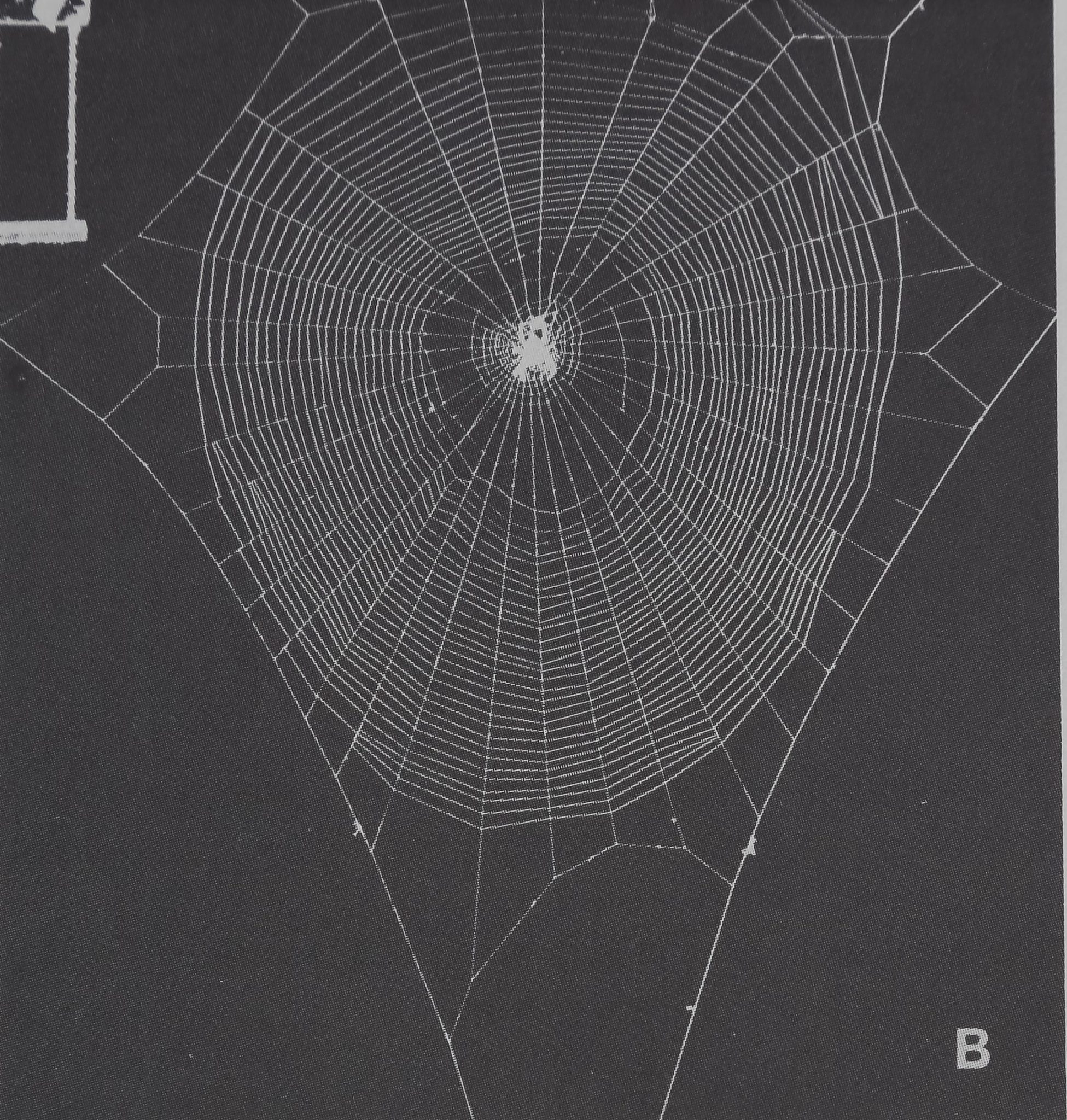
Fig. 3A-D. The webs of four different species built by just mature female spiders. A: Mature Araneus pima web. Spider’s description: age 230 days, body weight 320 mg, leg length 17 mm. Web measures: spiral area 67,407 mm2, center area 4669 mm2, thread length 24,190 mm, angle regularity 3.5°, mesh width 87.1 mm2. Note the irregular tangle of threads at the upper left corner of the frame. B: This web belongs to Araneus diade-mat us. Spider’s description: age 160 days, body weight 220 mg, leg length 17 mm. Web measures: spiral area 58,684 mm2, center area 5321 mm2, thread length 20,209 mm, angle regularity 2.4°, mesh width 77.8 mm2. Compare the shape of the web and the corresponding spiral area size to those for webs of Araneus pima and Neoscona domiciliorum. Compare spiral zone with that of a young web of the same species (Fig. 2B) in regard to thread regularity.


Fig. 3. Continued. C: Argiope aurantia web. Spider’s description: age 160 days, body weight 184 mg, leg length 23 mm. Web measures: spiral area 79,801 mm2, center area 9906 mm2, thread length 28,000 mm, angle regularity 3.8°, mesh width 100.6 mm2. Note the big spiral and center areas as well as the large mesh width compared to those for webs of the other three species. D: Neoscona domiciliorum. Spider’s description: age 135 days, body weight 100 mg, leg length 9 mm. Web measures: spiral area 60,307 mm2, center area 2442 mm2, thread length 27,230 mm, angle regularity 2.6°, mesh width 41.3 mm. Note the small center area and the irregularities in the spiral zone compared to those for a young web. Note that in Figs. 2 and 3 the spiders are sitting in the hub of the webs in their normal head-down waiting position. Compare the mature spiders’ webs with those of the young ones, as well as with data from Tables III, V, VII, and IX.
Quantitative Analysis of Orb Web Patterns in Four Species of Spiders 209


210
Risch
weeks immediately following the last molt. The three webs for each individual juvenile spider were selected at the period at which the spider was about midway between hatching and the final molt.
RESULTS
Measurements: Age, Body Weight, and Leg Length Results
Tables II and III illustrate that juvenile and just-mature spiders vary widely in age, body weight, and leg length. Only leg length in mature spiders shows no intrageneric ( Araneus)differences between species A. pima and A. diadematus; this is also true for comparisons between broods Aspl and Asdl. All other intrageneric comparisons between the two species show significant differences at both age levels for the three measurements. Even significant intraspecific differences are found, i.e., leg length between Aeal and Aea2 and between Nadi and Nad2 (Tables II and III). Similar intraspecific differences are found for body weight and age. Often no significant differences are found between species and broods for different genera, i.e., for leg length in Table II and among Aspl, Aeal, and Nad2.
Differences among genera, species, and broods are larger for mature spiders than for the young ones even when measured relative to within-brood variations (compare F values between Tables II and III). Means for age, body weight, and leg length in mature spiders (Table III) differ significantly from those^in juvenile spiders (Table II) in any comparison. Therefore, the t values for the paired samples are left out.
Discussion
One could assume that broods from the same species show similar age at midway point between hatching and maturity as well as after the last molt. That intraspecific differences deviate significantly is shown in Tables II and III for Araneus diadematus and Argiope aurantia broods.
The spiders were selected for analysis according to homogeneity in development, but at that point they are very heterogeneous in age, body weight, and leg length (Tables II and III). Reed and Witt (1972) found that even spiders from a single brood show considerable variation in body weight during the first months of life, and a brood can be divided into a significantly faster-growing, earlier-maturing group and a slower-growing, relatively late-maturing section; consequently, maturity for the members of one brood occurs over a wide range in time.
We may assume that differences in web patterns not only are due to differences between genera and species but also are influenced by the varia-
Quantitative Analysis of Orb Web Patterns in Four Species of Spiders
211
tion of age, body weight, and leg length. Some age-related changes are described by Witt et al. (1972); Ramousse (1973) as well as Benforado and Kistler (1973) showed that correlations exist between web measurements and body weight. A correlation between body weight and mesh width is described by Witt et al. (1972). I found in this study that mesh width is highly correlated with leg length. Since certain web measurements are correlated with age, body weight, and leg length, I also interpret web patterns in relation to these three variables.
Size Dimensions
The following four size dimensions are considered in this article:
1. Number of radii: Total number of radial threads in a web.
2. Spiral area: The ring around the center area delineated by the innermost and outermost spiral turn, expressed in square millimeters.
3. Center area: Includes the densely woven hub area and the inner free zone.
4. Thread length: Total computed viscid and nonviscid thread in a web, measured in millimeters.
Web size depends partly on metabolic activity, such as silk production for total amount of thread and energy conversion for the movements to distribute silk. These variables are also related to the rate of food intake and the rate of growth (Ramousse, 1973). They determine the width and fine structure of the area in which food can be trapped; and outdoors they are influenced by the space available, i.e., open area between supporting branches of a tree.
Results
Intrageneric comparisons in web size measures between young Araneus pima and A. diadematus show significant differences between species and broods for all four dimensions (Table IV). However, in many cases both Araneus species show no intergeneric differences in comparison with species or broods of Argiopeor Neoscona; i.e., spiral area between species Araneus diadematus and N. domiciliorum, center area between Araneus pima and N. domiciliorum, and thread length among broods Asd2, and Nadi, and Nad2 (compare Fig. 2 with data from Table IV).
In webs built by mature animals, the results for these variables (Table V) are opposite to those of webs from young spiders. In a comparison between Araneus pima and A. diadematus no significant differences are found, but both often differ significantly from other species. There are two exceptions: spiral area among A. pima, A. diadematus, and N. domicili-
Table II. Means (Je) and Standard Deviations (sd, based on individual web data) of Age, Body Weight, and Leg Length from Juvenile Spiders for
Genera, Species, and Broods®
| Araneus pima | Araneus diade mat us | Argiope aurantia | Neoscona domiciliorum | Differences
among |
|||||||
| Aspl | Asdl | Asd2 | Aeal | Aea2 | Nadi | Nad2 | |||||
| Number of spiders studied | 20 | 20 | 15 | 20 | 18 | 10 | 10 | ||||
| Number of webs studied | 60 | 60 | 45 | 60 | 54 | 30 | 30 | ||||
| Measurements | |||||||||||
| Age (days) | J 95.9 | 77.6 | 55.1 | 95.9 | 69.6 | 56.6 | 66.7 | ||||
| sd 21.6 | 29.6 | 17.0 | 14.7 | 23.4 | 13.9 | 12.4 | |||||
| Genera | |||||||||||
| A | A | B | F Si 10.636 | ||||||||
| Species | |||||||||||
| 5A | B | 5C | B | F * lg-O?6 | |||||||
| Brood | |||||||||||
| A | B | 5C | A | 5BD | CD | BC | F m: 15.53* | ||||
| Body weight (mg) | x 18.4 | 45.6 | 62.3 | 9.9 | 30.9 | 12.3 | 21.8 | ||||
| sd 12.5 | 30,2 | 36.6 | 8,7 | 26.4 | 10.2 | 14.9 | |||||
| Genera | |||||||||||
| A | B | B | F i 20.056 | ||||||||
A
B
A
A
Species F – 28.83d
Brood
5A B5 C AF 5D5 EF ADF F * 18.02*
Leg length (mm) x 6.7 9.2 9.9 6.6 8.3 5.2 6.9
sd 1.3 1.8 1.9 1.6 1.7 1.1 2.3
Genera
A B C F 1 16.66*
Species
AC B A C F = 22.67*
Brood
5A BC B 5A C5 5DS A55 Fm 14.15*
° The capital letters identify on the first line differences among genera, on the second line differences among species, and on the third line differences among broods. Means and standard deviations for genera and species are not shown. Two means do not differ significantly if they have at least one common letter, i.e., AB and AC, or A and A. Two means with no common letter but a number 5 differ significantly at the 5% level; they differ at the 1% level if no one letter is in common and no 5 is placed in the same position. F values indicate the variance among genera, species, and broods. Note that Araneus pima differs significantly on the 1% level from A. diadematus in every comparison and on both levels.
* Significant differences below P ÿ 0.01; analysis of variance.
Quantitative Analysis of Orb Web Patterns in Four Species of Spidei
Table III. Means and Standard Deviations of Age, Body Weight, and Leg Length for Mature Spiders’
| Differences | ||||||||
| Aspl | Asdl | Asd2 | Aeal | Aea2 | Nadi | Nad2 | among | |
| Number of spiders studied | 20 | 20 | 15 | 20 | 18 | 10 | 10 | |
| Number of webs studied | 60 | 60 | 45 | 60 | 54 | 30 | 30 |
| Measurements Mature spiders Age (days) | X
sd |
201.3
29.2 |
A | 159.9
40.4 |
116.7
28.9 |
201.5
22.7 |
A | 142.9
20.7 |
126.9
17.5 |
C | 141.9
14.3 |
Genera F = 13.61s | |
| A | B | C | B | Species F = 29.26s | |||||||||
| A | B | 5C | A | BD | CD | 5BD | Brood F = 25.59s | ||||||
| Body weight (mg) | X
sd |
260.8
39.6 |
A | 200.5
43.4 |
178.7
49.1 |
186.9
36.7 |
B | 164.6
26.7 |
66.7
23.9 |
C | 94.7
23.4 |
Genera F = 123.08s |
Leg length (mm)
Species
| A | B | B | C | F = 100.76“ | |||||||
| Brood | |||||||||||
| A | B | BD | 5B | 5CD | E | I | F = 52.31* | ||||
| X | 16.3 | 16.1 | 14.9 | 23.0 | 20.2 | 11.6 | 14.1 | ||||
| sd | 1.2 | A | 2.8 | 1.6 | 1.9 | B | 2.2 | 1.7 | C | 1.4 | Genera F = 161.99“ |
| Species | |||||||||||
| A | A | B | c | F = 108.61* | |||||||
Brood
5A AC 5BC D E F B F = 59.44* *
“For statistical explanations and letter order, see Table II. Leg lengths do not differ significantly between Araneus pima and A. diadematus. Ages between Asdl and Asd2, as well as between Aeal and Aea2, are significantly different.
* Significant differences below P = 0.01; analysis of variance.
Quantitative Analysis of Orb Web Patterns in Four Species of Spidei
Table IV. Four Dimensions in Size from Webs Built by Young Spiders Compared Among Three Genera, Four Species, and Seven Broods’
Differences
| Variables | Aspl | Asdl | Asd2 | Aeal | Aea2 | Nadi | Nad2 | among | |
| Number of radii x | 26,7 | 30,0 | 32.1 | 31,3 | 30,5 | 35,6 | 34,7 | ||
| sd | 3,5 | 4,6 | 4,5 | 6,1 | 5.3 | 6.6 | 3.4 | Genera | |
| 5A | m | C | F = 23.56* | ||||||
| Species | |||||||||
| A | B | B | C | F = 22.84* | |||||
| Brood | |||||||||
| A | B | BD | B | B | C | CD | F = 12.16* | ||
| Spiral area (mm2) x | 7,579 | 23,959 | 36,809 | 9,207 | 20,382 | 29,591 | 31,086 |
| sd | 4,018 | 14,122
A |
18,371 | 6,999 | B | 19,310 | 16,847 | C | 15,491 | Genera F – 16.46* |
| *A | B | 5C | B | Species Fm 30.91* | ||||||
| A | BD | CE | A | B | DE | CD | Brood F « 19.72* |
| Center area (mm*) | X
sd |
995
486 |
2,135
902 §A |
2,730
969 |
1,224
894 |
5A | 2,557
2,010 |
1,357
663 |
5B | 1,407
594 |
Genera F = 3.5 Ie |
| A | B | 5A | Species F – 17.00* | ||||||||
| A | 5B | C | A | BC | A | 5A | Brood F = 14.38* | ||||
| Thread length (mm) | X
sd |
4,867
1,638 |
12,667
6,041 A |
17,455
6,114 |
6,086
3,033 |
B | 9,752
6,660 |
17,078
8,050 |
C | 17,486
6,781 |
Genera F 1 37.28* |
| A | 5B | C | 5D | Species F m 51.04* | ||||||
| A | §B | c | A | 5D | C | C | Brood F = 28.96* |
a For statistical explanations, see Table II. Note the significant differences between species and broods from Araneus pima and A. diadematus for all four measurements. Both species show more similarities to species and broods from different genera than to their own genus.
* Significant differences below P * 0.01; analysis of variance, c Significant differences below P » 0.05; analysis’of variance.
Quantitative Analysis of Orb Web Patterns in Four Species of Spiders
Table V. Mature Spiders and Four Web Size Measures Compared Among Genera, Species, and Broods’
Differences
| Variables | Aspl | Asdl | Asd2 | Aeal | Aea2 | Nadi | Nad2 | among | |||
| Number of radii | X | 24.1 | 24.0 | 24.9 | 28.2 | 29.1 | 27.0 | 27.3 | |||
| sd | 3.8 | 2.6 | 3.3 | 3.1 | 3.8 | 3.2 | 3.3 | Genera | |||
| A | 5B | 5C | F = 30.27* | ||||||||
| Species | |||||||||||
| A | A | 5B | 5C | F = 20.576 | |||||||
| Brood | |||||||||||
| A | 5A | 5A | BC | b5 | 6C5s | 5BC5 | FM 10.02* | ||||
| Spiral area (mm2) | X | 51,006 | 45,792 | 44,293 | 71,200 | 79,366 | 44,088 | 49,153 | |||
| sd | 18,920 | 16,870 | 14,750 | 21,684 | 29,133 | 12,360 | 15,765 | Genera | |||
| A | B | A | F = 47.77* | ||||||||
| Species | |||||||||||
| A | A | B | A | F = 32.55* | |||||||
| Brood | |||||||||||
| A | A | A | B | B | A | A | F = 16.91** |
| Center area (mm2) | X
sd |
4,446
1,585 |
4,340
1,210 A |
4,361
1,027 |
10,414
2,910 |
B | 9,042
2,594 |
2,318
560 |
C | 2,768
772 |
Genera F = 237.72» |
| A | A | B | C | Species F = 158.50» | |||||||
| A | A | A | B | C | D | D | Brood F = 80.88» | ||||
| Thread length (mm) | X
sd |
17,783
6,986 |
16,920
5,131 5A |
17,315
4,879 |
22,262
6,145 |
B5 | 24,379
7,780 |
18,639
5,890 |
5C5 | 21,266
6,091 |
Genera F = 18.42» |
| AC | 5A | B5 | 5C5 | Species F = 12.36» | ||||||
| 5A | 5A | 5A | B5 | B | AC5 | 5BC | Brood F = 6.97* |
a Statistics are explained in Table II. Note that no significant differences exist within the genus Araneus between broods and species from A. pima and A. diadematus. Compare with Table IV. b Significant differences below P = 0.01; analysis of variance.
Quantitative Analysis of Orb Web Patterns in Four Species of Spiders 219
220
Risch
orum and thread length between A. pim and N. domiciliorum do not differ significantly. In comparing the data from spiral area (Table V) with the webs in Fig. 3 built by mature spiders, we find corresponding results: Argiope aurantia built the largest webs, followed by A. pima, N. domiciliorum, and A. diadematus.Argiope aurantia shows also the biggest central area; N. domiciliorum the smallest.
Argiope aurantia and N, domiciliorum differ in all web size measures significantly (Table V). Broods from Argiope and do not differ in
number of radii among the broods Aeal, Nadi, and Nad2; neither do they differ between Aea2 and Nad2 or in thread length among Aeal, Aea2, and Nad2, but they are significantly different in any other comparison in size (Table V),
In Table X, size dimensions are compared between immature (Table IV) and mature (Table V) spiders. Any comparison of the numbers of radii, except for brood Aea2, shows significant decrease with age. The spiral area, center area, and thread length increase in most cases significantly, with exception of thread length for Asd2 immature and Asd2 mature, which decreases slightly.
Discussion
The above results lead to the conclusion that size measures of webs built by immature spiders do not so much reflect directly their species-specific traits but rather reflect a spider’s age, body weight, and leg length, which may in turn show possible genetically determined components. Intrageneric comparison shows that Araneus pima differs significantly from A. diadematus in all web size dimensions. Table II shows that age, body weight, and leg length are significantly different between the two species. Even in comparing broods within the same species, Asdl and Asd2, the same trend is found. The two broods differ significantly in age and body weight (Table II), and they differ significantly in three of four measures in size (Table IV). Similar intraspecific differences may also be found in comparing the two broods from Argiope.
On the other hand, I find no significant differences between broods from different species and genera so long as they have similar age, body weight, and leg length; i.e., Aspl and Aeal do not show significant deviation in age, body weight, or leg length (Table II). Table IV shows that only the number of radii deviates, while the other three dimensions do not differ significantly.
The above results suggest that the mature spiders, in contrast to the young ones, built their web dimensions according to their species-specific traits rather than corresponding to age, body weight, or leg length. Following morphological classification, Araneus pima and A. diadematus belong
Quantitative Analysis of Orb Web Patterns in Four Species of Spiders
221
to the same genus. The same classification seems also to hold for their webbuilding behavior in regard to size dimensions. They do not differ in any comparison of size dimensions between either species or broods (Table V). However, there are significant differences in age, body weight, and leg length (except for leg length between Aspl and Asd2; see Table III).
In comparing dimensions in web size measures between immature and mature spiders (Table X), I find, as did Wiehle (1927) and Witt et (1972) for Araneus diadematus, a decrease in number or radii with increasing age for all species. Reed et al. (1969) recognized an increase in spiral area and thread length for Argiope aurantia between juvenile and mature spiders. This agrees with the increase in spiral area and thread length visualized in Tables IV, V, and X.
In size measures, I find that thread length varies more in the webs of young spiders than for matures. Center area varies extremely in mature spider webs. Fewer differences seem to exist in the spiral area for immature spiders than for matures. Number of radii varies on both levels similarly.
Dimensions of Regularity * The four regularity dimensions considered are defined as follows:
1. Angle regularity: Index for regularity of radii placement calculated
, as mean discrepancy of all adjacent central angles in a web.
2. Standard deviation of central angles: ;The following formula was used to compute this variable:
SD ÂTTT [N*21x)2]1/2
where N is the number of angles and xis the central angle size in degrees.
3. Standard error of median mesh size: This variable identifies irregular mesh departures from normal mesh distribution and is calculated by the following formula:
sEmed = –D-^11;253
where N is the number of meshes, X is the area of one mesh in mm2, and
1.253 is a constant factor.
4. Relative deviation of spiral turns S (south). This index may be .calculated for four directions (south, west, north, or east) and indicates the departure of spiral placement from the mean value. The following formula serves to compute this variable:
RD -SD/Z
222
Risch
where SD = \/(N – 1) x [A 2 X2 -(2 N is the number of spiral turns in any of the four directions, X is first differences between entries, and X = sum of first differences/[(number of entries) – 1],
Variables for regularity are measurements which indicate how precisely spiders move when they pull silk out of the spinnerets and attach it to other threads; it is an index for web-building precision.
Results
The F values for differences among genera, species, and broods for young spiders are much smaller for dimensions of regularity (Table VI) than for web size measures (Table IV); and fewer means are significantly different for regularity than for web size. No significant differences are found between Araneus pima and A. diadematus except for relative deviation of spiral turns S (Table VI). Both species show small intergeneric and interspecific deviation. In three generic comparisons, only one genus differs every time significantly. Likewise, in a three-species comparison, only one differs each time significantly. The smallest differences are found between broods.
Table VII shows that similar results for regularity are obtained for mature spiders; several even smaller differences are found, e.g., angle regularity within genera, species, and broods (Tables VI and VII), or standard error of median mesh size S among broods. For angle regularity, no intra-or intergeneric species or brood differences are found in webs from mature spiders.
Aspl, Asdl, Asd2, Nadi, and Nad2 (except for relative deviation of spiral turns S) show a decrease in regularity with aging in any of the four dimensions of regularity; for half of the comparisons, the differences are significant (Table X). The spiders of these five broods built less regular webs at the mature stage. Aea2 immature spider webs do not differ significantly in any regularity comparison with webs from the same brood of mature spiders (Table X). Only Aeal built significantly less regular webs as immatures than as matures in regard to angle regularity.
Discussion
Web regularity for juvenile (Table VI) and mature (Table VII) spiders, as considered in this study, does not seem to be genus, species, or brood specific; nor do the data show direct correlation with body weight or leg length. In comparing the results for regularity variables from young spiders with those from mature spiders (Table X), I assume that regularity is correlated with age rather than with a species-specific selective pressure or body
size.
Quantitative Analysis of Orb Web Patterns in Four Species of Spiders
223
Witt et al. (1972) also found that changes in web regularity are not a consequence of size of the spider but rather are correlated with age; i.e., in male and female webs of Araneus diadematus, regularity decreased signifi
cantly with age, while web size was correlated with sex and/or body size. It seems that regularity decreases during the lifetime (Table X); the spiders built in most cases less regular webs when mature. Comparing the picture of immature (Fig. 2) with that of mature (Fig. 3) spider webs, we again recognize that mature webs are less regular. It seems likely that regularity does not play an important role in the catching of flies, and decreases as a consequence of growing old.
Web Shape
To indicate web deviation from a circular form, the following three variables were calculated:
1. Width over length: This ratio expresses the oval shape of the web and is calculated by dividing the horizontal diameter of the catching area by the vertical diameter; it is 1.0 for a perfectly round web.
2. Radius north over south: By dividing the radius from hub to upper borderline of spiral turns by hub to lower limit, I obtain a measure of the vertical position of the hub in relation to the perimeter of the catching zone.
3. Radius east over west: The E/W index indicates the horizontal displacement of the hub independent of direction; in the computer program, the larger value was always recorded as the denominator.
Shape measurements of the orb show the departure of the web’s circular form, indicate the position of the hub in the spiral area, and are an expression of the up-down and right-left symmetry of the catching zone.
Results
Comparing webs’ width over length among genera by immature spiders (Table VIII), no significant differences are found. Larger differences exist at the species and brood level. Araneus diadematus differs significantly from A. pima for this variable, but not from Neoscona domiciliorum. Interbrood, intraspecies differences are found between Aeal and Aea2. All means for width/length are smaller than 1, indicating that the average web is oval shaped, with the long axis in the direction of gravity. The means for radius N/S and E/W indicate that no up-down or right-left symmetry exists for the webs under study.
For mature spiders, we also find that the average webs (Table IX) are oval shaped with the long axis in the southern direction. No significant dif-
Table VI. Four Regularity Measures from Juvenile Spiders Compared Within Genera, Species, and Broods®
| Variables | Aspl | Asdl | Asd2 | Aeal | Aea2 | Nadi | Nad2 | Differences
among |
|||
| Angle regularity (deg) | X | 3.9 | 3.6 | 3.6 | 4.9 | 4.2 | 3.3 | 2.9 | |||
| sd | 1.1 | 1.8 | 1.2 | 2.5 | 1.4 | 1.1 | 0.7 | ||||
| Genera | |||||||||||
| 5A | B | 5C | FB 17.296 | ||||||||
| Species | |||||||||||
| 5A | AD | 5B | CD | F =s 12.036 | |||||||
| Brood | |||||||||||
| AC | 5AE | . ACE^ | B5 | 5c|g. | af5 | DEF | F = 6.986 | ||||
| Standard deviation of center | X | 4.9 | 4.4 | 4.7 | 4.9 | 4.7 | 3.8 | 3.6 | |||
| angles (deg) | sd | 1.4 | 1.3 | 1.6 | 2.0 | 1.6 | 1.0 | 0.7 |
| A | A | B | Genera F‘& 10.52# | ||||||||
| A | A | A | B | Species F = 7.89’’ | |||||||
| A | 5AB | A5 | A | A5 | BC5 | 5C | Brood F = 4.25* |
| Standard error of median mesh size S (mm2) | 3c
sd |
0.056
0.033 |
0.044
0.048 5A |
0.035
0.043 |
0.055
0.071 |
5B | 0.133
0.347 |
0.017
0.015 |
A | 0.017
0.009 |
Genera F = 6.18» | |
| AB | A | B | A | Species F = 4.26* | ||||||||
| A | A | A | A | B | A | A | Brood F = 3.58» | |||||
| Relative deviation of spiral turns S | X
sd |
0.258
0.064 |
0.368
0.155 A |
0.336
0.088 |
0.385
0.107 |
B | 0.378
0.125 |
0.396
0.146 |
BC | 0.357
0.090 |
Genera
wM 10.81* |
| A | B | B | B | Species F = 15.29* | ||||||
| A | BC | 5B | 5C | BC | 5C | BC | Brood F = 8.25* |
a For statistical approach, see Table II. Note the smaller intra- and interdifferences among genera, species, and broods than in Table II, especially for standard error of median mesh size S. Among broods, only Aea2 differs significantly from the others.
* Significant differences below P 0.01; analysis of variance.
Quantitative Analysis of Orb Web Patterns in Four Species of Spiders 225
| Table VII. Four Regularity Measures Compared Within Genera, Species, and Broods in | Webs Built by Mature Spiders® | ||||||||
| Differences | |||||||||
| Variables | Aspl | Asdl | Asd2 | Aeal | Aea2 | Nadi | Nad2 | among | |
| Angle regularity (deg) x | 4.8 | 4.2 | 4.7 | 3.9 | 4.0 | 4.0 | 4.1 | ||
| sd | 2.1 | 1.4 | 1.6 | 1.5 | 1.7 | 0.9 | 1.4 | Genera | |
| A | A | A | F = 2.94 | ||||||
| Species | |||||||||
| A | A | A | A | F = 2.37 | |||||
| Brood | |||||||||
| A | A | A | A | A | A | A | F= 1.42 | ||
| Standard deviation of center x | 5.8 | 5.4 | 5.8 | 4.8 | 4.5 | 4.8 | 4.9 | ||
| angles (deg) sd | 1.7 | 1.2 | 1.4 | 1.2 | 1.6 | 1.0 | 1.3 | Genera | |
| A | B | B | F = 11.67* | ||||||||
| 8A | 5A | B | 5B | Species F1 7.78* | |||||||
| 6AC | 5AC | A5 | 5B | B | BC5 | BC5 | Brood F = 4.34* |
*P*!H 9ZZ
| Standard error of median mesh size S (mm2) | X
sd |
0.119
0.101 |
5A | 0.096
0.070 |
0.089
0.095 |
0.156
0.158 |
A | 0.116
0.191 |
0.053
0.048 |
5B | 0.043
0.042 |
Genera F = 6.13* |
| 5AB | AC5 | B5 | 5C | Species F = 4.43* | ||||||||
| 5AB | AC5 | ac5 | Bs5 | ABC | ac5 | 5C5 | Brood F = 2.55c | |||||
| Relative deviation of spiral turns S | X
sd |
0.431
0.141 |
A | 0.377
0.103 |
0.356
0.099 |
0.496
0.150 |
B | 0.401
0.097 |
0.478
0.125 |
AB | 0.345
0.062 |
Genera FI 5.98* |
| A | B | A | AB | Species F = 6.50» | ||||||
| 5AD5 | 5B | B | C5 | ab5 | cd5 | B | Brood F = 7.60* |
a For data evaluation, see Table II. Note the small F values and mean differences among genera, species, and broods in all four measurements. Angle regularity does not deviate significantly in any comparison.
* Significant differences below pH 0.01; analysis of variance. c Significant differences below P= 0.05; analysis of variance.
Quantitative Analysis of Orb Web Patterns in Four Species of Spiders
Table VIII. Three Dimensions in Shape and One in Fine Structure (mesh width) Compared Within Genera, Species, and Broods at the Spiders’
Juvenile Period0
| Variables | Aspl | Asdl | Asd2 | Aeal | Aea2 | Nadi | Nad2 | Differences
among |
|||
| Width/length | X | 0.88 | 0.79 | 0.76 | 0.86 | 0.79 | 0.81 | 0.84 | |||
| sd | 0.10 | 0.12 | 0.10 | 0.19 | 0.16 | 0.11 | 0.07 | ||||
| Genera | |||||||||||
| A | A | A | F% 0.29 | ||||||||
| Species | |||||||||||
| 5A | B5 | SC5 | 5BC | F = 6.736 | |||||||
| Brood | |||||||||||
| 5A | BC | B5 | AD | BC | 5BDC | AC5 | FM 4.92b | ||||
| Radius ratio N/S | X | 0.69 | 0.68 | 0.73 | 0.85 | 0.87 | 0.78 | 0.79 |
| sd | 0.13 | 5A | 0.23 | 0.17 | 0.26 | B5 | 0.22 | 0.16 | 5C5 | 0.11 | Genera F = I6.62b |
| 5A | 6A | B5 | 5C5 | Species F = 11.10* | |||||||
| 5ACD | 5AC | ACD | B | B | 5BC | 5BD | Brood F k 5.77* |
228 Risch
| Radius ratio E/W | X | 0.85 | 0.81 | 0.72 | 0.89 | 0.94 | 0.88 | 0.86 | ||||
| sd | 0.12 | 0.15 | 0.16 | 0.17 | 0.15 | 0.11 | 0.09 | Genera | ||||
| A | B | B | F = 18.80* | |||||||||
| Species | ||||||||||||
| 5A | B | 5C | AC | F = 15.84* | ||||||||
| Brood | ||||||||||||
| AC | 5A | B | CD | D5 | 5CD | AC5 | Fl 10.16* | |||||
| Mesh width (mm2) | X | 21.9 | ■’ 3.2jlf | 40.4 | 19.2 | 34.8 | 23.9 | 24.7 | ||||
| sd | 8.9 | 13.3 | 17.3 | 12.0 | 21.3 | 11.5 | 8.3 | Genera | ||||
| 5A | AC | 5BC | F 1 3.74c |
| A | B | A | A | Species F ~ 10.556 | ||||||
| A | 5BD5 | 5C | A | bc5 | A5 | ad5 | Brood F= 10.15* |
a For data evaluation, see Table II. Note that for width/length no significant differences exist among genera (F = 0.29). Mesh width does not deviate significantly among the following broods: Aspl, Aeal, Nadi, and Nad2. bSignificant differences below P m 0.01; analysis of variance. c Significant differences below P – 0.0$; analysis of variance.
Quantitative Analysis of Orb Web Patterns in Four Species of Spiders 229
K>
O
Table IX. Three Shape Measures and One Variable in Fine Structure (mesh width) for Mature Spiders®
| Variables | Aspl | Asdl | Asd2 | Aeal | Aea2 | Nadi | Nad2 | Differences
among |
||
| Width/length | X | 0.89 | 0.84 | 0.83 | 0.90 | 0.88 | 0.91 | 0.85 | ||
| sd | 0.13 | 0.10 | 0.09 | 0.17 | 0.14 | 0.17 | 0.10 | |||
| Genera | ||||||||||
| A | A | ’ A | Fl 1.59 |
| A | A | A | A | Species F = 2.63 | ||||||
| A | A | A | A | A | A | A | Brood F = 1.97 |
| Radius ratio N/S | X | 0.70 | 0.57 | 0.64 | 0.80 | 0.98 | 0.81 | 0.83 | ||||
| sd | 0.13 | A | 0.14 | 0.18 | 0.18 | B | 0.22 | 0.13 | B | 0.15 | Genera F = 51.29* | |
| A | B | C | C | Species F = 36.84* | ||||||||
| 5A | B | AB | 5C | D | 5C | C | Brood F = 22.76» |
| Radius ratio E/W | X
sd |
0.88
0.15 |
A | 0.75
0.17 |
0.70
0.17 |
0.88
0.12 |
B | 0.93
0.13 |
0.90
0.10 |
B | 0.87
0.10 |
Genera F = 22.01* |
| Species | ||||||||||||
| A | B | A | A | F = 25.91* | ||||||||
| Brood | ||||||||||||
| A | B | B | A | A | A | A | F = 13.87» | |||||
| Mesh width (mm2) | 3c | 88.1 | 78.8 | 72.4 | 111.1 | 106.6 | 54.9 | 52.4 | ||||
| sd | 22.9 | 19.6 | 19.9 | 25.4 | 26.0 | 16.1 | 13.1 | |||||
| Genera | ||||||||||||
| A | B | C | F = 76.91* |
| 5A | B | C | D | Species F = 53.49 | ||||||
| A | AB | 5B | C | C | 5D | D | Brood F – 27.09* |
«
a Statistical evaluation is explained in Table II. For width/length, no significant differences were found for any comparison on the genus, species, or brood level. Mesh width for Aspl does not differ significantly from that for brood Asdl but deviates from that for Asd2 and any other intergeneric broods.
* Significant differences below P = 0.05; analysis of variance.
Quantitative Analysis of Orb Web Patterns in Four Species of Spiders
232
Risch
ferences for this shape measure are found in any comparisons among genera, species, or broods. The shape index variables (radius ratio N/S and E/W) indicate that the mature spiders neither build circular webs nor locate the hub in the center.
The means for width/length, compared between immature (Table VIII) and mature (Table IX) spiders, show that webs built by mature spiders are less oval shaped; four of seven comparisons indicate a significant deviation in this regard (Table X). Juvenile as well as mature spiders seem to localize the hub in the same place. As for the horizontal and vertical hub position, only three of 14 comparisons differ significantly: Asdl differs for radius ratio N/S and E/W, Aea2 in the vertical placement of the hub (Table X).
Discussion
Juvenile as well as mature spiders build oval-shaped webs. This oval web form does not seem to be characteristic for certain genera or species. Only small differences are found between species and broods for immature spiders (Table VIII), but no significant deviation among genera; for mature spiders, no significant differences for width over length are found (Table IX).
If the web’s oval shape resulted entirely from the effect of gravity (Mayer, 1953), the old, heavy spider should build relatively long webs; there was sufficient space in the cages for that. My results show the opposite. The Spearman rank correlation coefficients were calculated for body weight and width over length for young ( rs = 0.786; significant differences below P = 0.05) and mature spiders (rs = 0.214; no significant differences below P -0.05). The oval shape of mature spiders’ webs did not show as great an effect due to weight; adults built more circular webs than young ones, even though adults were many times heavier.
The vertical displacement of the hub from the center seems to be more related to specific spider groups than width over length. Especially in mature spiders (Table IX) the radius ratio N/S seems to be typical for certain species or broods. Araneus pima differs significantly from A. dia-dematus; both are also significantly different from Argiope aurantia and N. domiciliorum. Because of the way in which the computer program is written, the exact horizontal position of the hub is not identified; only the degree of hub displacement is calculated. Between juvenile and mature spiders a significant horizontal eccentricity exists only for brood Asdl (Table X). The horizontal hub position in webs from adult A. diadematus seems to be different from that for the other species (Table IX). In general, the shape of the web built by young and mature spiders does seem to be similar across genera or species.
Table X. Significant Differences (indicated by asterisks) for Each Variable Between Single Immature and Mature
Spiders Within Broods0,6
| Variables | Aspl | Asdl | Asd2 | Aeal | Aea2 | Nadi | Nad2 | Differences between juvenile and mature spiders |
| Number of radii | ** | ** | ** | ** | ** | ** | F = 165.Il6(I) | |
| Spiral area (mm2) | ** | ** | — | ** | ** | * | ** | F = 242.196 (I) |
| Center area (mm2) | ** | * * | ** | ** | ** | * | F = 513.41 (I) | |
| Thread length (mm) | ** | ** | ** | ** | — | ‘ — | F = 108.996 (I) | |
| Angle regularity (deg) | * | 1 Rl 1 | * | ** | . —_ | * | F =, 6.306 (I) | |
| Standard deviation of center angles (deg) | ** | ** | * | ** | F = 21.96* (I) | |||
| Standard error of median | * | — | ** | mmm | F = 15.836 (NI) | |||
| mesh size S (mm2) | ||||||||
| Relative deviation of spiral | ** | — | — | ** | — | ** | F = 50.156 (I) | |
| turns S | ||||||||
| Width/length | _ | * | * | ** | ** | — ■ | F = 20.54″ (NI) | |
| Radius ratio N/S | — | ** | — | ** | — | — | F = 0.72 (I) | |
| Radius ratio E/W | . -i/•’ : | * | — | — | — | F = 0.52″ (NI) | ||
| Mesh width (mm2) | ** | ** | ** | ** | ** | ** | ** | F = 1086.98″ (I) |
°The comparisons were done by means of a t test for paired samples (Snedecor and Cochran, 1972). F values result from global comparisons for each variable between young and adult spiders. I, Interaction between ages and broods exists; NI, no interactions were found. Note the increase in web size measures for most of the broods (Tables IV and V), the decrease for regularity (Tables VI and VII), and the constancy in upper-lower and right-left symmetry (Tables VIII and IX). Mesh width increases greatly with age for any brood (Tables VIII and IX).
6 Significant differences below (**) Pm 0.01 and (*) P |f0.05; analysis of variance.
Quantitative Analysis of Orb Web Patterns in Four Species of Spiders 233
234
Risch
Web Fine Structure
To elucidate the web’s fine structure (the density or distribution of the threads in the spiral zone), the mesh width was calculated by the following formula:
, . 11 spiral area
Mesh width = —-———|——
number of radii x mean number of spirals
The formula shows that mesh width measures a complex process of movements which include spacing of radii and laying of viscid spirals. Radius construction requires determination of central angles; other movements are required for laying sticky spiral.
Results
Differences in mesh width between juvenile spider webs on the generic level are very small. Only Araneus and Neoscona differ at the 5% level. In comparing species, we see that only Araneus diadematus is significantly different in mesh width from the other species. Intraspecific deviations are found between the two broods Asdl and Asd2, as well as between Aeal and Aea2. Broods show more intergeneric and interspecific relationships. Aspl does not differ significantly in mesh width from the broods Aeal, Nadi, and Nad2.
For mature spiders, mesh width deviates significantly among genera, species, and broods from different species (excluding Aspl and Asdl). Table X shows significant larger mesh width for webs built by mature spiders in all broods.
Discussion
Mesh width determines prey size. It may be the result of contingencies determined by body measures like leg length, which in turn may have been selected according to prey size.
In this study, mesh width is directly correlated with leg length. A Spearman rank correlation coefficient was calculated for immatures ( =
0.857; significant correlation at the 5% level) and adults (r, = 0.965; significant correlation at the 1% level) for mesh width and leg length. The rs values show significant correlations at both levels. Spiders with the longest legs (Tables II and III) built webs with the largest mesh width (Tables VIII and IX); e.g., mature spiders from brood Aeal (Table III) have the longest legs (23.0 cm) and Nadi the second shortest (11.6 cm). Similarly, Aeal (Table IX) has the largest mesh width and Nad2 the smallest. The range in leg length (Table III) between broods corresponds directly to the range in mesh width (Table IX) with the exception of Nad2.
Quantitative Analysis of Orb Web Patterns in Four Species of Spiders
235
GENERAL DISCUSSION
In the study of species-specific or genus-specific behavior, the behavior should be significantly complex to permit analysis of many details, and it should be accessible to recording and measuring. It is advantageous to select an animal with a highly patterned behavioral output; the experience of the individual as well as the environment in which it behaves should play a minimal role in the patterned movements of the organism. Mayr (1974) defines such a system as “closed,” explaining that the neuronal connections which underlie the execution of the behavior pattern are largely determined by the genetic code, rather than by conditions under which the individual grows and develops. There is much evidence to indicate that spider webbuilding is such a closed system.
Reed et al. (1970) discuss experiments where spiders were raised under extreme conditions, so that they either could not build any web (narrow tube) or never caught a fly in their web (mouth-feeding). In comparing webs built by these spiders at adulthood with those of littermates which had built and used webs daily, no differences were found in size, regularity, shape, and fine structure, indicating that web construction is based neither on practice nor on reinforcement.
In additionp my investigations were performed on spider webs which were built under extremely uniform environmental conditions: all animals were raised in the same laboratory with controlled temperature and light programs. All animals were fed one housefly twice a week so that at least food supply was uniform. All built in aluminum frames of equal size and shape sufficiently large to avoid limitations through narrow supports. This eliminated the influence of such factors as place of birth, growing conditions, and nature of prey. Therefore, differences in webs of different species are largely correlated with a species-specific genetic code, which is responsible for the development of the neural circuits, which in turn guide webbuilding. My finding that species of one genus can be distinguished by their web pattern confirms the above conclusion.
Two of my species belong to one genus, while two others belong to different genera. I assume that species of the same genus are more closely related than species of different genera; similarities and differences in morphological characteristics of the animals have been used to classify them as to the proximity of such relationship. If closed behavior patterns are as characteristic for a species as body surface, one should find web patterns of different but closely related detail in the species of one genus, and less closely related web patterns in different genera. My measurements support this assumption, because the size measures of the webs of mature spiders are more closely related within the same genus ( than between
genera. Since the species ( A.pima and A. diadematus) of the same genus
236
Risch
(Araneus) occupy different habitats (Levi, 1971) and build similar webs, one could assume that web-building is related to the gene pool background.
Certain details of web structure may be important for species isolation in mating. The mature male spider approaches the female by means of the web. By the male’s drumming on peripheral threads of the female’s web, the long courtship starts. The vibration and tension changes in the web are recognized by the female; communications between female and male may begin. It seems obvious that at this point species and maturity recognition of the other sex would be advantageous. Species-specific characteristics of the web, such as size of spiral and center area, shape, and stabilimentum, could modulate the pattern and extent of vibration (Langer, 1969) in such a way that the male recognizes the appropriateness of its courtship behavior. In this study, not only species-specific features are found, but also specific web characteristics for single broods (i.e., littermates of single cocoons); such differences are always less than those between species, but this weakens the argument for the possible role of web pattern in species isolation. However, there is strong evidence that vibration, if important in courtship and mating of web builders, is only one of the ways of communication between male and female; Blanke (1974) and others discuss evidence that females and webs of araneids are releasers of pheromones, which apparently play an important role.
If the web’s resonant properties are important for a successful mating within the same species, one could assume that mature spiders build highly species-specific webs. For this reason, I suppose that webs from mature spiders deviate in certain web details not only from webs of other species but also from the webs built during immaturity. In this study, a large discrepancy was found for most variables of size measures and fine structure between immature and mature spiders, mature webs showing more species specificity. Therefore, the adult web is not an enlarged projection of a young spider’s web; it is rather a specialized outcome with species-specific features.
The spider’s orb web is a highly evolved construction and a result of a long evolutionary process, but one wonders whether it is the product of a monophyletic development. Because no orb web fossils have been found, interpretation of the evolution of orb webs must be highly speculative. One may assume that since the orb web was “invented,” natural selection affected genes controlling the nervous system, fixing specific details in web structure and spider behavior for the survival of the individual and species. In order to be effective for a species’ survival, orb webs had to adapt to environmental changes. Certain orb web structures were changed more drastically than others; some structures could have been lost or added (e.g., stabilimentum). Presumably, constant adaptation in regularity and shape of the web was not directly necessary for the survival of the species, even in a
Quantitative Analysis of Orb Web Patterns in Four Species of Spiders
237
continually changing environment. On the other hand, measures of size and fine structure play an important role for species survival; for this reason, I suspect modifications in these web details, according to environmental changes in time and space. In this study, we find that webs from mature spiders of different genera, species, or broods seem to be very similar in regard to their web measures in regularity and shape; for angle regularity and width/length, no significant differences were found. Significant differences were found in comparing dimensions in size and fine structure.
On a highly speculative basis, one could imagine that our four species had common ancestors and are an expression of a monophyletic line. Presumably, continuous adaptation of web regularity and shape was not directly necessary for the survival of the species, even if the environment was continously changing. In contrast, the orb web characteristics of size and mesh width were important for the trapping function; changes had to occur for these web details in order to be effective in species survival.
There are species of spiders with short legs which build rather large, wide-meshed webs (i.e., Hyptiotes paradoxus; Peters, 1953) and other species where the animals grow to large size but persist in building narrow-meshed webs ( Nephilaclavipes). In this experiment, I found that leg length
is directly correlated with mesh width at both age levels. Peters’ findings have only limited applicability to my findings; he compared spiders with extremely different web-building behavior, very distant family relationship, and extremes in body characteristics. Of the four species I studied, all show similar web-building behavior; in these species, mesh size could be the result of leg length or vice versa.
In this study, only a group of 12 different web parameters was selected. Many more web details could be considered for web-building behavior studies. Species-specific structures such as frame area, stabilimentum, and irregular tangle of threads (see Fig. 2A) were not considered. Statistical computations not mentioned here showed that 15 additional calculated parameters would not change our results significantly.
Of the thousands of existing spider species, the four selected are only a small sample. Since the variation in every web parameter within species, broods, and individuals is very great, it appears wise to deal first with a small number of species, but with many individuals.
ACKNOWLEDGMENTS
I would like to thank Dr. P. N. Witt for his assistance during all stages and gratefully acknowledge the assistance of Mrs. M. Scarboro for all laboratory work and Mrs. R. Daniels for administrative help. My thanks are also due Dr. J. O. Rawlings, with whom the statistical evaluations were discussed.
REFERENCES
238
Risch
Benforado, J., and Kistler, K. H. (1973). Growth of the orb-weaver Araneus diadematus, and correlation with web measurements. Psyche 80:90-100.
Berman, J. D., and Levi, H. W. (1971). The orb weaver genus Neoscona in North America. Bull Mus. Comp. Zool 141:465-500.
Blanke, R. (1974). Das Sexualverhalten der Gattung Cyrtophora als Hilfsmittel fiir phylogenetische Aussagen. Proc. 6th Int. Arachn. Congr. 1974, pp. 116-119.
Committee on Form and Style, Conference of Biological Editors (1964). Style Manual for Biological Journals, American Institute of Biological Sciences, Washington, D.C., pp. 1-117.
Grasshoff, M. (1968). Morphologische Kriterien als Ausdruck von Artgrenzen bei Rad-netzspinnen der Subfamilie Araneinae. Abh. Senckenberg. Naturforsch. Ges. 516:1-100.
Kaston, B. J. (1964). The evolution of spider webs. Am. Zoologist 4:191-207.
Kullmann, E. (1958). Beobachtung des Netzbaues und Beitraege zur Biologie von Cyrtophora citricola Forskal. Zool. Jb. Syst. 86:181-216.
Langer, R. M. (1969). Elementary physics and spider webs. Am. Zoologist 9:81-89.
Levi, H. W. (1968). The spider genera Gea and Argiope in America. Bull. Mus. Comp. Zool. 136:319-352.
Levi, H. W. (1971). The diadematus group of the orb-weaver genus Araneus north of Mexico. Bull. Mus. Comp. Zool. 141:131-179.
Mayer, G. (1953). Untersuchungen ueber Herstellung und Struktur des Radnetzes von Aranea diadema und Zilla x-notata mit besonderer Beruecksichtigung des Unterschiedes von Jugend- und Altersnetzen. Z. Tierpsychol. 9:337-362.
Mayr, E. (1974). Behavior programs and evolutionary strategies. Am. Scientist 62:650-659.
Peters, H. M. (1939). Problème des Kreutzspinnen-Netzes. Z. Morphol. Oekol. Tiere 36:179-266.
Peters, H. M. (1953). Weitere Untersuchungen ueber den strukturellen Aufbau des Radnetzes der Spinnen. Z. Naturforsch. 8b:355-370.
Peters, P. J. (1970). Orb web construction: Interaction of spider {Araneus diadematus Cl.) and thread configuration. Anim. Behav. 18:478-484.
Ramousse, R. (1973). Body, web-building and feeding characteristics of males of the spider Araneus diadematus. Psyche 80:22-47.
Reed, C. F., and Witt, P. N. (1972). Growth rate and longevity in two species of orb-weaving spiders (Araneae: Argiopidae). Bull Br. Arach. Soc. 2:111-112.
Reed, C. F., Witt, P. N., and Scarboro, M. B. (1969). The orb web during the life of Argiope aurantia (Lucas). Dev. Psychobiol. 2:120-129.
Reed, C. F., Witt, P. N., Scarboro, M. B., and Peakall, D. B. (1970). Experience and the orb web. Dev. Psychobiol. 3:251-265.
Snedecor, G. W., and Cochran, W. G. (1972). Statistical Methods, Iowa State University Press, Iowa City, pp. 1-593.
Szlep, R. (1966). Evolution of the web spinning activities: The web spinning in Titanoeca albomaculata Luc. (Araneae, Amaurobidae). Israel J. Zool. 15: 83-88.
Wiehle, H. (1927). Beitraege zur Kenntnis des Radnetzbaues der Epeiriden, Tetragnathiden und Uloboriden. Z. Morphol. Oekol. Tiere 8:468-537.
Wiehle, H. (1929). Weitere Beitraege zur Biologie der Araneen, insbesondere zur Kenntnis des Radnetzbaues. Z. Morphol. Oekol. Tiere 15:262-308.
Witt, P. N. (1971). Instructions for working with web-building spiders in the laboratory. BioScience 21:24-25.
Witt, P. N., Reed, C. F., and Peakall, D. B. (1968). A Spider s Web, Springer-Verlag, New York, pp. 1-107.
Witt, P. N., Rawlings, J. O., and Reed, C. F. (1972). Ontogeny of web-building behavior in two orb-weaving spiders. Am. Zoologist 12:445-454.

Quantitative Analysis of Orb Web Patterns in Four Species of Spiders Behavior Genetics
Dr Peter Witt‘s work applied drugs to orb web spiders using spider web geometry to identify biochemical abnormalities. Peter Witt (1940’s – 1990’s)’s comprehensive body of work features articles (Life, National Geographic), Spider Communication, Life on a Thread and his work with the North Carolina Department of Mental Health.
After what he always described as his accidental discovery of the effects of drugs on the geometry of the orb web, a major portion of Dr Peter Witt’s subsequent written work (over 100 papers and 3 books) was concerned with the behavior and biology of spiders. Scientific Articles offer Peter Witt’s complete works, while human interest articles include features like experiments by NASA in the Skylab with the orb web spider in the early 1970’s.
“He was a man of courage when it was dangerous to be so, of boldness in his experiments and of generosity with his colleagues, students and friends” said Charles Reed in Peter Witt’s 1998 obituary. He is survived by his daughters, Elise Witt and Mary Witt.