Images Collection


View this article in Search Friendly Plain Text
NOTE: This plain text article interpretation has been digitally created by OCR software to estimate the article text, to help both users and search engines find relevant article content. To read the actual article text, view or download the PDF above.
Reprinted from Science, September 10, 1965, Vol. 149, No. 3689, pages 1190-1197
Spider-Web Building
Measurement of web geometry identifies components in
a complex invertebrate behavior pattern.
Peter N. Witt and Charles F. Reed
The different species of orb-weaving spiders spin webs with different characteristic patterns, although variation occurs from day to day and from individual to individual within a species. Building the orb requires a complex sequence of actions by the animal. This sequence may be disturbed by various means without halting construction altogether; a web of some sort is built even after profound physiological interferences: under certain drug conditions, with legs missing, and with any of a variety of lesions in the central nervous system. Changes *in the form of the web are more or less specific to particular physiological conditions; some elements of the usual pattern may be disturbed while others are not.
This circumstance is useful in pharmacological investigation, for the distinctive effects of drugs on web geometry provide a basis for classifying and distinguishing among drugs. The survey of drug effects upon web patterns presented below will illustrate the use of the web for the detection and differentiation of drugs.
Inevitably the question arises as to what alterations in bodily function are reflected by the changes in pattern. The web is a convenient and precise record of the operation of the sensory and motor apparatus of the spider. If the web could be selectively deranged, elementary functional systems could presumably be related to particular pattern characteristics. We may view the spider as a web-building machine which integrates sensory information regarding the current state of muscles, spinning glands, appetite, and other
Dr. Witt is associate professor of pharmacology at the Upstate Medical Center of the State University of New York, Syracuse. Dr. Reed is associate professor of psychology at Temple University, Philadelphia, Pennsylvania.
bodily conditions before and as it builds its web. As will be seen, faulty information can be supplied in one way or another, with consequent distortion of pattern.
We are presently attempting to identify the elementary functions involved in web construction, together with the stimuli which ‘’trigger” and guide them. Certain kinds of information become incorporated into the design; evidence of the nature of this information has emerged progressively
from investigations of the effects of
^ .
drugs, from other types of experimental interference with function, and from extension and refinement of analysis of the pattern. Our review of these inquiries will emphasize the behavioral record which the web comprises and the functions implied by this record. Ability to duplicate or simulate web-building, for instance with a computer program, promises an even more precise criterion for deciding whether those functions have been adequately identified.
Procurement of Webs
Whether we try to define the geometry of the orb built by an undisturbed animal, or whether we interfere with building in specific ways in order to evaluate the consequences, the method is basically the same. We make the webs visible by spraying them with white paint, photograph the sprayed webs, and measure and evaluate the patterns in terms of plane geometry. Because so many factors influence daily web-building, many webs have to be evaluated in order to obtain statistically valid data.
The experimentally desirable achievement of obtaining one web per spider every day all year round in the labora
tory is forestalled by a number of problems. Inadequacy of control sometimes becomes apparent only belatedly, to cast doubt on findings previously attributed to the experimental variable. For example, a systematic difference in webs built in the laboratory on Mondays eluded detection for some time before it became evident that, unless a web was destroyed by the end of the day, it was probable that the spider would eat it and as a result build a larger web on the next day (/). This discovery led to the simple procedure of not using Monday webs as controls in experiments. Other problems are less amenable to solution.
Some web changes are associated with maturity of the spiders (2). The simplest solution would seem to be experimentation with mature animals. However, there is really no “mature” web: after the female has reached full size, other factors, such as egg development, affect weight and hence web pattern, and males build too infrequently in later life to be experimentally useful. During winter months, the mortality rate of spiders is high and weaving is relatively infrequent, even in the warm laboratory. The low frequency of weaving can be attributed in part to the relatively even temperature in the laboratory’ and to brevity of daylight (5). The matter has been improved somewhat by installing a programmer which controls the temperature, providing a temperature minimum in the early morning hours, followed bv a steep rise, and by providing artificial illumination which simulates 16 hours of daylight.
Egg- cocoons can be hatched all year round but. for reasons not yet clear, only relatively few cocoons release spiderlings in the laboratory. About 200 animals can be obtained from one cocoon. Except for the tedious job of feeding all the spider-lings, raising them on a diet of fruit flies and w;ater presents no difficulties. It is interesting, although experimentally inconvenient, that even “litter-mates” from one cocoon, raised under similar conditions, grow at different rates.
Frequency of web-building varies with the species: female Zygiella-x~ notata Cl. and Arcineus diadematus Cl. build daily, writh interruptions during molting; the former can be easily caught out of doors in central Europe, the latter in Europe and the northern part of the United States. While
l
Zygiella lives for only about 8 months, A. diadematus can survive for 18 months. Araneus sericatus Cl., although convenient in that it can be caught out of doors nearly all year round and builds a web very similar to that of A. diadematus, spins less frequently, about every other day.
Obtaining the Record
The webs, which are built in 20 to 30 minutes in the early morning hours, are fragile and hardly visible. For behavioral analysis moving pictures of web-building in progress are unsatisfactory because of the low photographic contrast of the thread, to say nothing of the expense and labor of accumulating and analyzing films (4). Homann’s method [see Peters, Witt, and Wolff (5)] of placing cases with completed webs into a box filled with smoke of ammonium chloride was an early solution to the problem of the visibility of the orb; we find it easiest to remove the spider and coat the threads lightly with Krylon white glossy spray paint.
The web is photographed with 35-millimeter high-contrast film. So that each thread will be reproduced as a thin white line contrasting sharply with the background, the case is placed in front of a black box and lighted from both sides. A weight, composed of a lead bar and three cotton threads 20 millimeters apart, is suspended in the plane of the web to serve as a scale for enlargement, as well as an indicator of the direction of gravity (as in Figs. 1 and 2, for example).
The record is thus obtained cheaply and stored readily. The experimenter may return to it in order to measure selected aspects of the pattern: data on drug effects were gathered in this manner.
As will be described later, current evaluation of the photographs consists essentially of three steps: first, measurements are made of selected points; then components of the pattern are calculated from the figures; and finally, statistical comparisons are performed. The first two steps, which require enormous numbers of calculations, have particularly been facilitated by the use of a computer (4, 6).
Effects of Drugs
Experiments with drugs first demonstrated the selective consequences of changing the physiological condition of the spider. It is of course instructive to interfere with the building process itself, for example by destroying radii as they are placed. The various phases of construction, such as the changeover from nonviscid to viscid thread, have been investigated in this manner (see 7 and 8 and Fig. 1).
Drug experiments have been performed with web-building spiders for various reasons: the web test has been used to distinguish between drugs when their effects have been indistinguishable by methods testing the behavior of other animal and human subjects (5, 9-16). For example, it has been possible to differentiate between ways in which the stimulants caffeine and amphetamine act (11), or between the depressant actions of chlorpromazine and of barbiturates (17).
In other experiments the same drug was used as a tool to compare orb-web construction in different species (5, 18). There is evidence that spiral placement after administration of metham-

Fig. 1 (left). An uncompleted web. The spider was interrupted during web-building after it had just started to construct the sticky spiral at the right and bottom of the web; the preliminary spiral is still intact. At this point in the construction a change in the movement pattern and thread material occurs. Fig. 2 (right), complete web of an adult female Araneus diadematus Cl. (about 5 months old). The original distance between two of each of the three parallel lines in the upper right corner was 20 mm; the lines indicate the direction of gravity (see text). Twenty-four radii connect the densely woven hub with the frame threads; the catching spiral covers an oval ring between the center and frame areas.
2
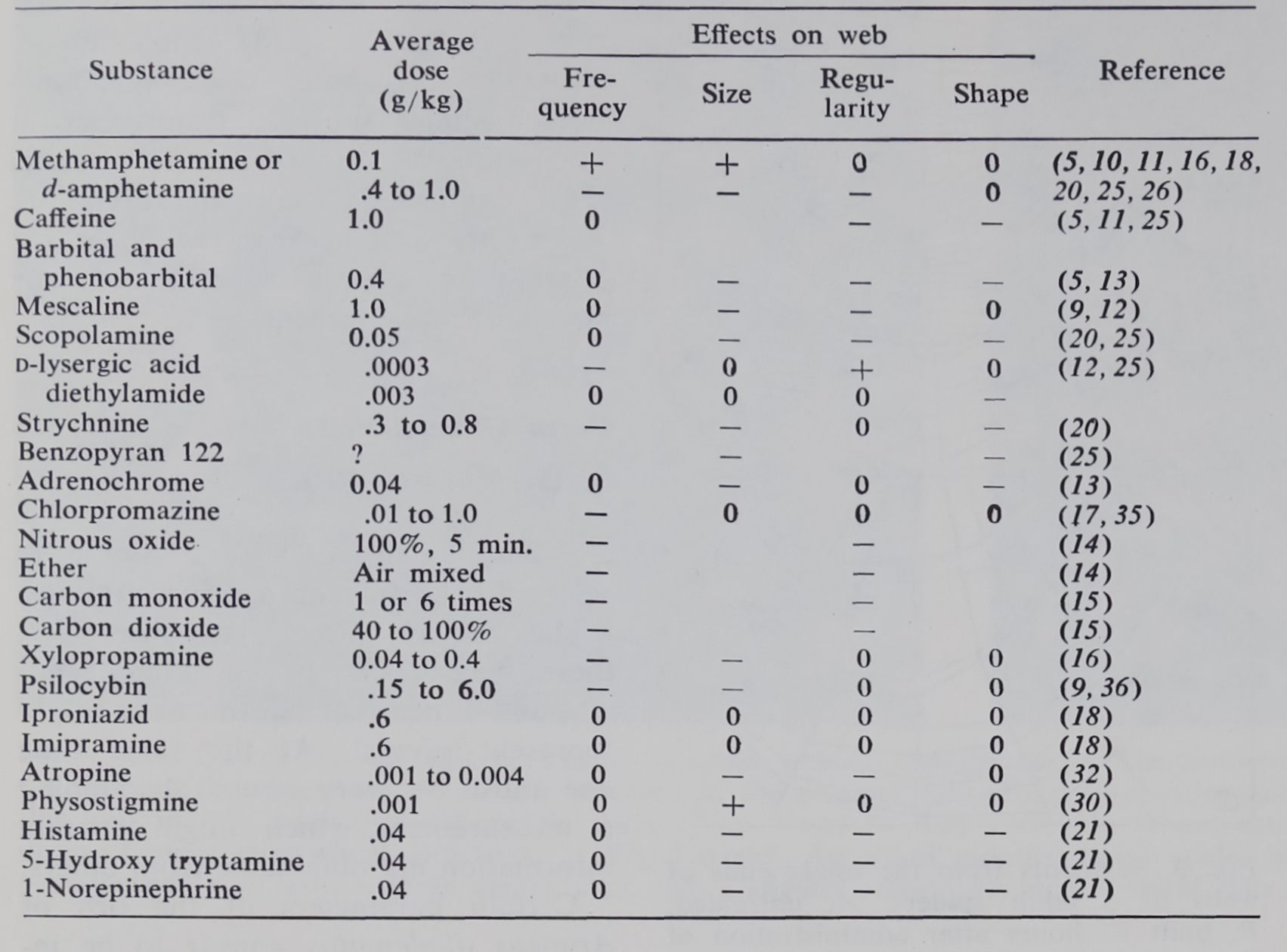
Table 1. Effects of drugs on spiders’ web-building behavior. For the purpose of uniformity the following simplifications are made: the drug name, not the salt used, is listed; drug combinations are left out; route of application is not given; effects of anesthesic or lethal doses are deleted; no distinction among spider species is made. Absence of an entry signifies that the particular parameter was not measured. Plus refers to web-building more frequent than controls, larger or more regular webs; minus refers to a decrease in frequency, size, or regularity and to a shape diffrent from control; zero indicates no effect. Significance is set at the 1-percent level of probability.
phetamine, for instance, shows the same irregularities in the webs of Zygiella (10), Araneus (17), and Nephila (19).
Drug combinations were tried out to measure interaction and possible potentiation (3, 18). It was found, for instance, that, a low dose of rf-amphetamine changed web-building significantly when the spider had been previously treated with a low dose of iproniazid, each substance alone being ineffective in the amounts given. This potentiation coincided with a change in amphetamine metabolism in the body, resulting in high concentrations of amphetamine in iproniazid-pretreated spiders.
Different routes of application, for
instance, injection of drugs as compared with oral administration (18, 20) or with inhalation (14, 15), did not produce measurably different effects. In the latter experiments the case was placed in the gas mixture just after the spider had finished building the radii; spiral construction was severely disturbed. One subtance, carbon monoxide, was applied repeatedly to find out whether the sensitive web test would indicate a lasting effect of “chronic poisoning” with the gas. No such influence was found.
Table 1 shows a simplified summary of results with a variety of drugs. Not all data were obtained in the same way; the species of spider, type of web (young or old), and evaluation
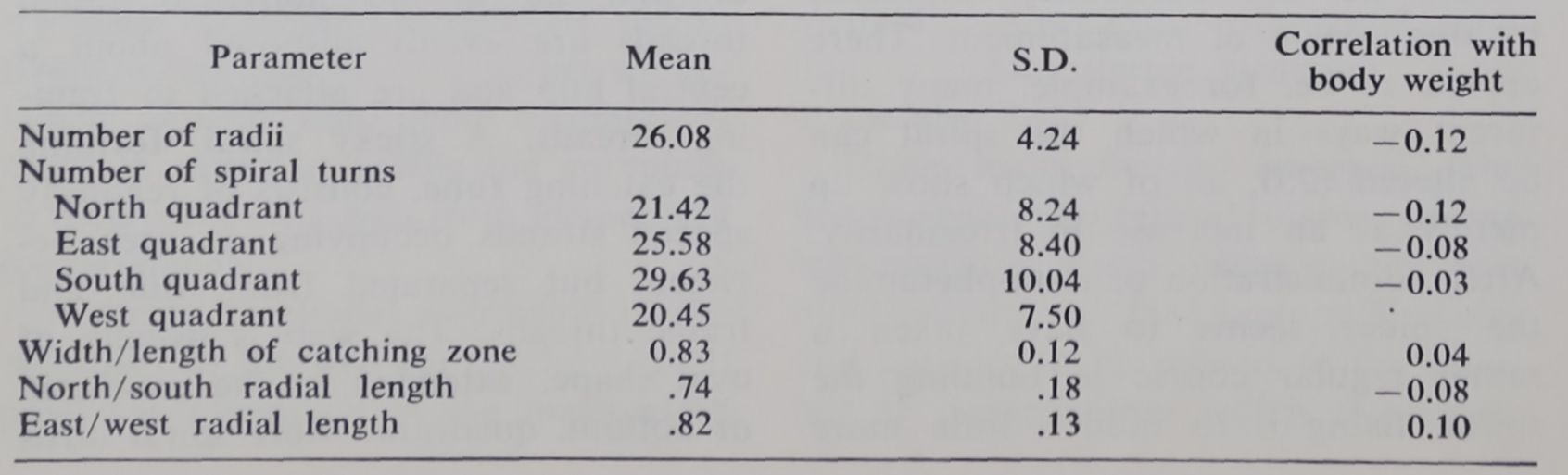
Table 2. Values for those web dimensions which are not dependent upon weight (age) of spider. They are based on a sample of 103 female Araneus diadematus. Note that shape and number of elements are not significantly correlated with body weight.
varied. However, an overall result seems clear: the web test reveals effects of many of the drugs tried and can be used to distinguish among their effects.
Efforts were made to interpret changes in web parameters in terms of alterations in animal behavior. An example is the effect of the muscle-relaxant tranquilizer diazepam (Valium), given in a dose of 100 milligrams per kilogram of body weight 24 hours before web-building time. A decrease in regularity of the spiral above and below the hub was observed, with no change on the right or left. This can be interpreted in terms of movements of the spider as it builds the spiral in different parts of the orb; on top and bottom it shifts its body horizontally across vertically running radii, whereas at the sides it lets itself down vertically or climbs straight up. This differential spiral effect may provide a useful test for muscle-relaxant drug effects.
With the use of the information of the sensitivity of web-building to pharmacological agents, the web test was applied to the search for foreign substances in the body fluids of mental patients (21-24). One problem in these studies was the choice of human controls, because urine, plasma, or cerebrospinal fluid of the control subjects may contain many substances which affect the web test. Follow-up experiments seem necessary in the cases where body fluids were found effective (27, 22).
Extending the Pattern Analysis
The drug studies show that specific combinations of changes, rather than individual modifications in web geometry, are important. It is necessary, therefore, to examine as many aspects of the pattern as possible and ultimately to define the basic constituents of the record. Those basic constituents are not necessarily synonymous with the elements of the web considered as a geometric figure alone. For instance, the data for diazepam suggested that different operations were required for placement of the horizontal and vertical spiral threads. The spider is not a draftsman; spirals, or for that matter any structure in two different quadrants of the web, are not necessarily equivalent.
Given relatively few index measurements, a computer can reconstruct
3


almost any aspect of the web pattern we choose. The problem was the selection of those index measures. Our procedure is as follows:
Step 1: The 35-millimeter film is projected with the help of an enlarger to original web size. Measures are derived by application of a polar coordinate system. Using an arbitrary point in the hub as center, radii are identified by their compass direction, spirals by their position along the radii. It was found sufficient to measure the distance from the web center for each spiral turn only along the radii near the four cardinal directions; for all other radii, only the distance of the innermost spiral turn, of the outermost spiral turn, and of the frame thread were recorded. These figures, together with identifying data, were fed into an IBM 1620 computer, in punched card form, and stored in its memory.
Step 2: The computer is programmed to combine the figures in such a way that three kinds of measurements of web characteristics can be derived:
1) Size measures: total length of thread, number of radii, number of spiral turns in each of the four directions, and .extent of catching area (that which is covered by the sticky spiral), center area, and frame area.
2) Shape: defined by the ratios of the north to the south and the east to the west radii, and of the vertical to the horizontal diameters. A quotient between each of two areas is also calculated.
3) Regularity measures: obtained for the radial angles and spiral spacing: the mean of all differences between adjacent angles is a convenient expression of central angle regularity. A radial thread may* be thought of as being divided into short segments by successive spiral turns. The sizes of the segments vary. By subtracting the lengths of adjacent segments from each other, and by calculating the mean of all such differences, an expression for spiral regularity is derived. This expression can be calculated separately for the horizontal and vertical directions.
Step 3: With the present computer program (4) each component in up to 20 webs can be compared to the respective component in 20 other webs with the /-test or the chi-square method. Such comparision has been performed in two ways: either all webs built by one spider before an experiment have been compared with all those

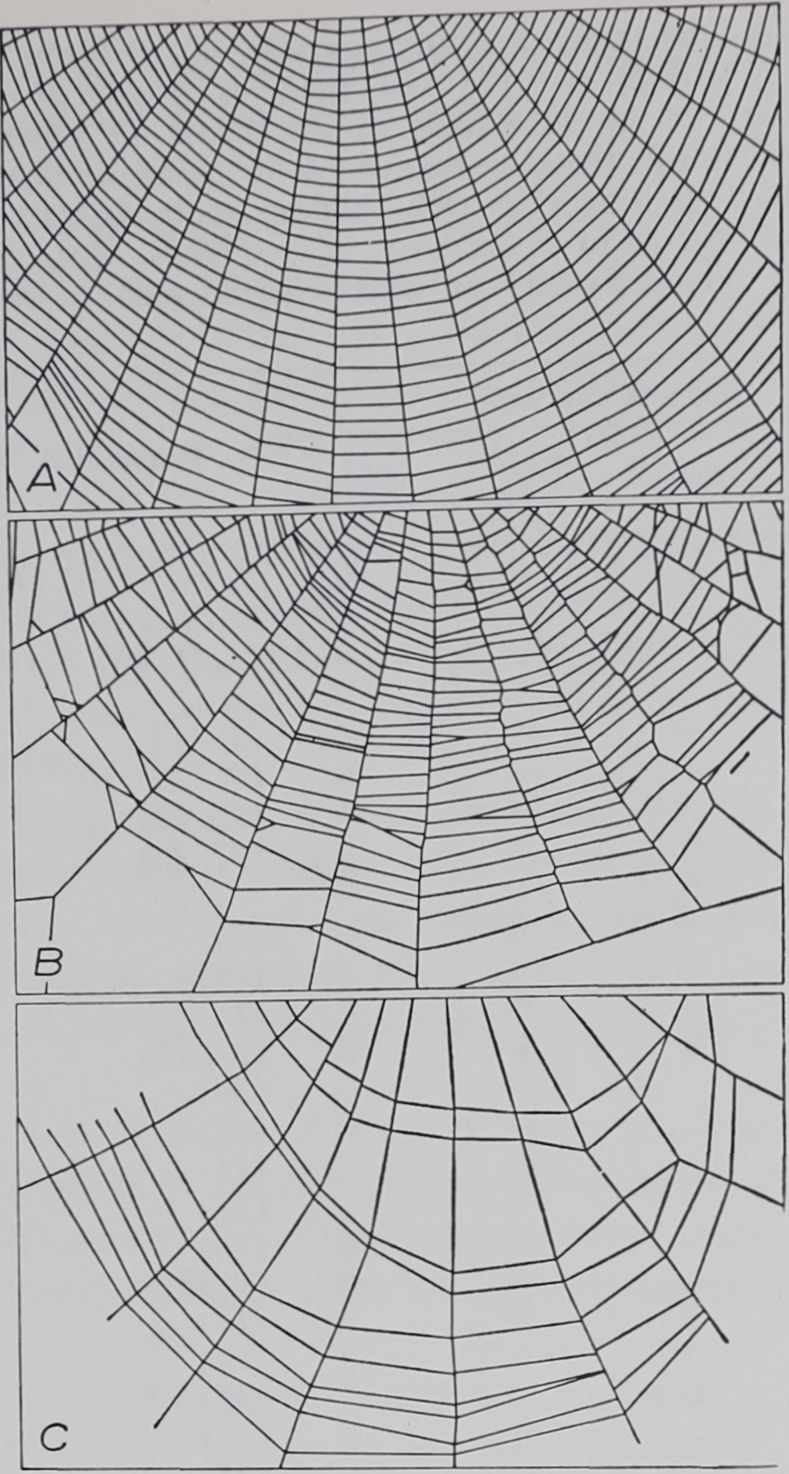
Fig. 3. Segments from the lower ends of webs of 3 adult spiders. A, untreated; B, built 12 hours after administration of ^-amphetamine (0.6 g/kg); C, built 12 hours after administration of scopolamine (0.05 g/kg). The figures point out a limitation of our web evaluation method: although the types of disturbances in the animal’s movements in B and C were clearly different, the computer evaluation with the present program would merely indicate in both webs a decrease in spiral regularity.
built afterwards, or a group of webs built by many spiders on a control day has been compared with webs of the same animals on experimental days. The first method lends itself more to the establishment of the effect of a lesion; the latter is used to measure changes brought about by drug application. However, the data which the computer prints out in table form can be combined in many other ways if additional information is desired.
Web changes have been observed which are not adequately expressed by our system of measurement. There appear to be, for example, many different ways in which the spiral can be altered (25), all of which show up merely as an increase in irregularity. After administration of //-amphetamine the spider seems to have taken a rather regular course in building the spiral, fixing it to radii a little more
to the right or to the left than in the webs of controls. After administration of scopolamine, by contrast, the path of the spiral-laying spider goes farther and farther off course (Fig.3). A tentative analysis postulates a defect in fastening but not in orientation in the case of the amphetamine-treated spider, whereas the scopolamine-treated spider shows a disturbance in orientation. A motion-picture analysis (26) confirmed such interpretation. There are instances in which direct observation proves superior to our system of measurement.
Form Characteristics of the Normal Web
While dimensions of the web were selected originally on a priori grounds, it has always been recognized that these measures may be interdependent, and hence that one measure might represent several. At the same time one must be wary about abandoning a measurement which might contain information not obtainable from others.
Certain parameters of the orb of Araneus diadematus appear to be independent of the age or, more particularly, the weight of the spider. They maintain essentially the same values for the webs of young, light animals and for those of old, heavy spiders with a lifetime of web-building behind them. The characteristic form of the web may be appreciated from an examination of Figs. 4 and 5 and Table 2.
The structural components of the web are built in the following sequence (see also Figs. 1 and 2): (i) a horizontal bridge on which the whole web is suspended; (ii) a Y-structure determining the center: (iii) radii and frame threads; (iv) a temporary spiral, curving in three to four wide turns from center to periphery; and (v) a sticky or catching spiral running in tight pendulum and circular turns from the outside toward the hub.
In the completed web of the female (Fig. 2) at least after the first month of life, 20 or 30 nonviscid radial threads are evenly disposed about a central hub and are attached to framing threads. A sticky spiral, forming the catching zone, consists of regularly spaced strands occupying an area between, but separated from, hub and frame threads. The web is usually of oval shape, extended in the southern, or bottom, quadrant. More spiral turns
4

Fig. 4 (left). One of the first webs of a newly hatched Araneus diadematus Cl. The two white lines, parts of a metal measure resting on the floor of the cage, indicate 20 mm distance in the original web. Note similarities and differences in details of pattern between this web and the adult and old spiders’ webs (Figs. 3 and 5). The very thin thread can be recognized only by means of the fine drops of spray paint which stick to it. Fig. 5 (right). An old (8 months) heavy spider builds with a relatively short, thick thread a wide-meshed web with a large center area and a slightly irregularly spaced spiral. The scale in this figure is the same as in Fig. 2.
are found here than in other parts, but greater variation is also observed. However, the overall shape of the web and the number of threads in a sector remain relatively constant and independent of the growth of the spider.
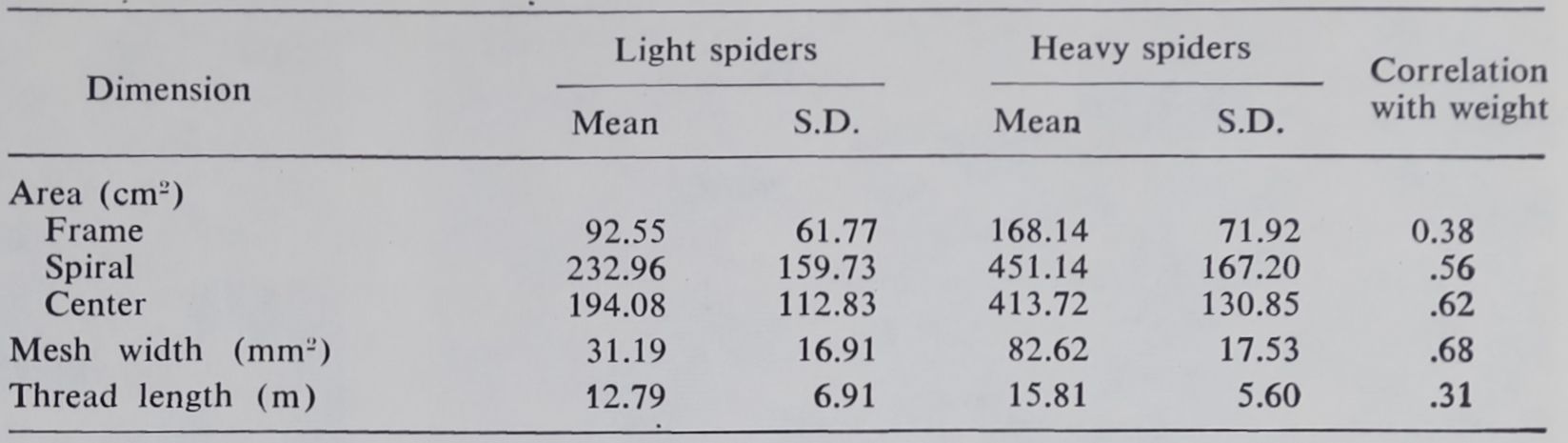
On the other hand, some web measures show significant variation with the weight of the spider (Table 3). The total space occupied by the web is one of these: three zones may be distinguished within it. The center area consists of the hub and the free zone, the annular region in which no viscid spiral is found. The spiral area composes the catching zone of the web, where sticky spirals are fastened to the radii. Both of these regions increase in size as the spider’s body weight increases. A less marked relationship is found between weight and framing zone. This portion of the web, which is irregular and variable, lies outside the catching area. The area of the framing zone is difficult to measure definitively in any case, but the values we obtained depend also upon assumptions made in order to enable the computer to reconstruct the web from elementary measurements.
A direct functional expression of the efficiency of the web for catching small prey was attempted in the mesh-width
measure. This is in effect an overall measure of the fineness of partitioning of the space occupied by the web. Body weight of the spider is positively correlated with mesh size.
Finally the total length of the thread used, including both structural and viscid elements, shows a significant but relatively slight correlation with body weight. Figuratively, as the spider grows, she throws her net more widely, but a net not provided with material to insure always the same density. The small value of the correlation may be due to the fact that the measure of length expresses only one aspect of the thread, which is known to vary also, for example, in thickness (9).
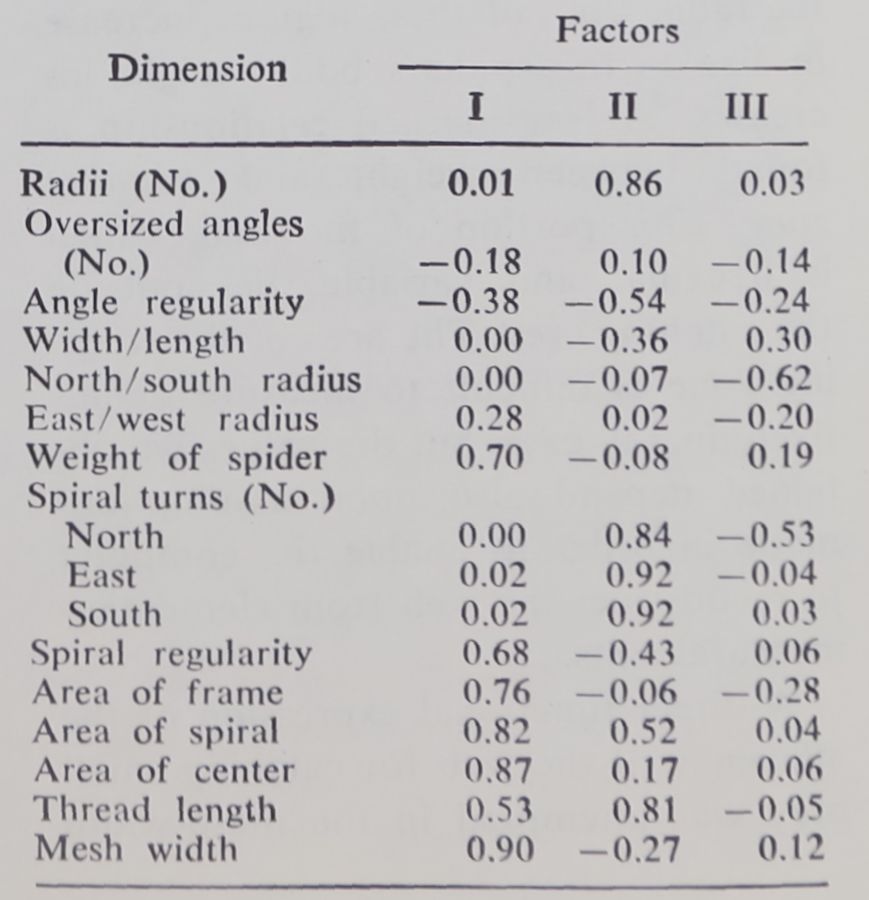
Factor analysis provides a further procedure for assessing the independence of a priori measures of characteristics of the normal web. A matrix of intercorrelations among 16 such measures was analyzed by the centroid method, followed by orthogonal rotation to obtain simple structure (Table 4; see also 27).
Three main factors emerged. The first is apparently related to area: among our measures mesh width is the best representative of that factor. The second appears to be related to the number of portions into which the catch
ing zone is divided: a possibly equivalent measure may be simply a count of the trapezoid spaces in the spiral area. The third factor is a less distinct one and has something to do with the asymmetry of the web, that is, the position of the hub with respect to the geometric center. None of our measures seems to be a simple representative of this factor.
Our expressions of spiral and angle regularity were found in the factor analysis to contain an artifactual element we had not previously detected: webs with relatively few radii or spirals show a spurious degree of irregularity.
Thus analysis of the measures reveals certain redundancies and interdependencies. It is desirable, of course, to reduce redundancies and to measure parameters which are as independent as possible. Functional correlations—depending upon the operations of the web builder—are sought, rather than logical correlations. Although the expressions for spiral and angle regularity which we have used heretofore are not independent of web size and number of threads, they are not merely alternative expressions for these components. For example, the correlation found between regularity and area breaks down in spiders with random
5
lesions in the cephalothorax (28). Such lesions generally lead to a reduction in web size, but webs of 13 out of 35 spiders showed a significant change in spiral regularity without decrease in area.
The Plan
Certain aspects of web construction can be associated with normal and abnormal conditions of the spider’s body.
It has been found that alterations in weight, rate of thread synthesis, and leg size and function are accompanied by changes in building techniques and consequently in the dimensions and pattern of the web. We will examine some of these conditions and their relation to web architecture; each is more or less essential to the execution of some component of the final structure.
Weight and leg length. For instance, as the sample of normal spiders in Table 2 indicates, heavier animals build webs of larger area. (The discussion which follows refers especially to the area covered by the spiral.) In the course of its life, however, a spider reaches a stage beyond which increase in weight is not accompanied by an increase in web size. Changes continue to occur in thread and detail, but the area does not exceed an apparent maximum except perhaps under certain drug conditions.
The sequence of events is as follows (see 2): During the first 8 to 10 weeks of life, the young spiders grow rapidly in weight and in leg length (the latter through successive molts); the web area also expands, and the web contains an increasing number of radii and spiral turns. In the following 4 months the mesh size increases; that is, the number of radial and spiral threads declines while the web area remains relatively constant. In the same period, body weight doubles but leg length, like web size, does not change. (See Figs. 4 and 5 for representative webs of young and old spiders, respectively.)
Since leg length and web size follow a parallel course, it would seem that the length of the leg somehow determines the extent of the space to be occupied by the web, at least under laboratory conditions, where the physical features for support remain constant. Weight, while it yields place to leg length as the basic determinant of web area, is essentially related to mesh size. That relationship is clarified by
Table 3. Values for those web dimensions which vary with weight (age) of the spider. Measurements were made for single webs for 51 females weighing not more than 100 mg (range, 7 to 100 mg) and 29 females weighing 150 mg or more (range, 150 to 500 mg). Correlations are based on a sample of 103 female spiders of all weights. Observe that web size is a significant constituent of all measures which positively vary with weight.
considering further the webs made by young and old spiders.
Not only do old spiders build webs with wide meshes, but their threads are thicker than those of young animals. Pursuing the hypothesis that weight rather than maturation accounts for differences in thread thickness, we made young spiders heavy by attaching lead weights to their abdomens (9). They thereupon built webs with thick thread and wide meshes. Thus increase in weight somehow requires a change in the characteristics and distribution of thread.
The course of events may be hypothetically reconstructed as follows: with increase in body weight, a thicker thread is required for support; this accommodation probably occurs as the spider hangs on its thread in the first stages of web construction. There are two other limiting conditions: the supply of silk and an apparent requirement to maintain a web of an area proportionate to the size of the animal (probably mediated through performance of legs). A compromise is reached by altering the density of the threads so that mesh size is increased. This arrangement would at first appear to be contrary to the interest of the spider, allowing prey to slip through more readily than before. However, preservation of web size permits catching of an undiminished number of large prey and such loss as occurs is certainly less catastrophic than building no web at all—which would he the consequence if the thread were too thin to support the animal. Further, the loss is perhaps less than might at first appear. The web detains rather than imprisons (29) prey; the sticky thread of the spiral will impede an insect long enough for the spider to seize it.
It is interesting that, whereas mesh size increases in the first web built after addition of weight, the mesh returns to its previous size only gradual
ly (in the next 3 days) after the weight is removed (9). The latter adjustment presumably is made in the face of less urgent circumstances than the initial one. Be that as it may, weight imposes certain requirements which have to be accommodated to other conditions and requirements, such as those set by leg length and by a variable to which we now turn for further examination, the supply of silk.
Thread supply. Early in the drug studies it was observed that changes in the size of webs and in the number of radii and spirals were associated with different kinds of drugs of a wide variety. Stimulation or inhibition of thread-protein synthesis was suspected to be the significant factor.
The total amount of silk in a web could be estimated on the assumption that thread protein contained a constant amount of nitrogen. Consequently whole webs were digested in selenium sulfuric acid and the nitrogen in the digested material was determined (/, 9). Following the observation that treatment with the cholinergic drug phy-sostigmine produced larger webs which contained 33 percent more protein than
Table 4. Factor loadings after rotation.


did controls (30), Peakall tested the effect of physostigmine, carbachol, and a number of other cholinergic substances on speed of incorporation of labeled amino acids into silk protein (37). For intact spiders, as well as for isolated glands in vitro, these drugs increased speed of thread production. It was shown later (32) that atropine, an anticholinergic, significantly slowed the process. A cholinergic regulation of silk synthesis, possibly through neural routes, was postulated. Consequently, the large, dense webs produced after administration of physostigmine and the wide-meshed webs produced after administration of atropine reflect stimulation and inhibition of silk production, respectively.
A relation between thread supply and size of web might have been suspected a priori, but the consequences of thread supply for web pattern are less apparent. A number of possibilities come readily to mind when one considers how the amount of material available might be manifested in web form. The spider could continue to work until it ran out of thread, with the pattern showing varying degrees of incompleteness or extravagance, depending upon supply. On the other hand, work might halt when the demand of other variables was satisfied, even if the thread-producing glands were not emptied; in this case one would expect. the pattern to be preserved unless there was a drastic shortage of silk. How does relative abundance of thread material regulate web-building behavior? Two general phenomena have to be described in attempting to answer this question.
First of all, there is evidence that all or most of the available silk material is utilized in a web. Peakall found the ampullate glands (which supply the basic structural thread) and the aggregate glands (furnishing the sticky material for the spiral) to be virtually empty at the completion of web-build-ing (37).
If more material is available, the area of the web is made larger. In addition to the effect of physostigmine already cited, we may consider the consequences of allowing the spider to eat it$ daily web rather than removing the threads from the cage (7). When this is done for a number of days, the successive webs become larger and denser and have thicker threads than under control conditions. Presumably this development is a consequence of abundance of material.
If on the other hand, the supply of
material is diminished, radii and spiral turns decrease in number and the threads used for them become thin, and ultimately the size of the web is also reduced. At least this course of events can be observed in spiders deprived of food for extensive periods. With less and less silk available, the web undergoes necessary changes. Food-deprived spiders also become lighter in weight, apparently using body constituents for silk synthesis (33).
The second phenomenon to be considered is that the spider does not simply run out of material and thereupon give up web-building. There seems to be a definite “planned” endpoint. Information concerning the amount of material remaining in the glands appears to be integrated into the process of building.
This point may be supported by two lines of evidence.
1) In the final stages of construction the spider pauses after completion of scaffolding (frame, hub, radial threads) and the temporary spiral (Fig. 1). It then begins to work its way in from the periphery of the web, attaching the sticky spiral to each radial thread as it comes to it. If the animal stopped only when it ran out of material, the central area, with its characteristic free zone and hub, would be variable with respect to spiral area. Instead the sizes of the spiral and central zones are highly correlated (r = 0.80 in 103 webs). We infer that the spider starts and halts the spiral at specific points in the pattern, probably guided by a central representation of amount of silk remaining in the glands. Depletion of thread material and completion of pattern converge upon a precise end-point.
2) Related evidence is supplied by experiments of Koenig (3), in which radii were destroyed as they were constructed. The spider produced new radii until it had furnished half again as many as would be expected in its normal web. It then began the process of building the spiral, attaching viscid thread to the inadequate scaffolding. Apparently the construction of the spiral was initiated when the spider’s thread supply was reduced to a minimum required for the spiral, this condition preempting all others.
In summary, thread material is distributed in the workspace according to supply but also in accordance with other requirements. Those requirements are such as to preserve (i) area (at an optimum size for the particular stage of development the spider has reached)
and (ii) the characteristic overall geometric design or “plan” of the web.
Measuring function of the legs. An essential element in the realization of the web plan is accuracy of leg movement. In addition to carrying the spider about and seizing prey, the legs are essential in testing distances. The spacing of radial and spiral threads appears to require certain maneuvers. Probing movements of the first legs can be observed in the process of building, and removal of those legs affects pattern. Webs built by spiders deprived of one leg or of two ipsilateral legs have grossly irregular central angles. The spiral is also abnormal in a manner reminiscent of webs built under the influence of amphetamine (Fig. 3).
Conclusions
Certain physical and physiological conditions current at the time of web-building seem to constitute elementary information which is incorporated into the form of the web of the orb-weaving spider Araneus diadematus Cl. The amount of thread material available, length of leg, weight of the body, and various locomotor and measuring functions apparently enter into a complex integration which determines the form of the “well-planned” web. Preservation of size, regularity and shape of the pattern are prominent in a hierarchy of requirements.
At present it is not possible to state whether some events are only derivative of other basic variables and circumstances encountered by the spider in the course of web construction. Body weight, for example, may determine speed of movement and, rate of extrusion of thread, or may be of significance only as a parameter of information transmitted b^ afferents from the legs. Some events, for example rate of expenditure of thread material, seem to require central integration at the start of building, while others, such as radial and spiral separation, are immediate responses to sensory information.
In the terminology of the computer, one may inquire into the form of the program in existence at the start of construction, and the nature of the data processed then and during web-build-ing. The behavioral records so conveniently provided by the spider are instances of more or less successful runs of the central program. It is not difficult to imagine computer simulation of web-building behavior, although’ the
7
basic elements required for such simulation are only in the process of being identified.
It is tempting to find in the investigation of the webs of the orb-weavers a solution for the problem of studying animal behavior with the methodological stringency of the laboratory but without restricting or oversimplifying the behavioral repertory {34). Unlike most laboratory animals, Araneus need be taught nothing nor be in any way impeded in its pursuit of survival. In its own good time it produces a record of a significant portion of its behavioral capacity, in a form which is readily measured and tested. Of course, mating and manner of seizing and devouring prey are behavioral events which cannot be studied by web evaluation. In other traditional areas of behavioral concern, such as the operation of sensory and learning functions, the restricted adaptive capacities and the highly specialized sensory range of the invertebrate spider may be advantages rather than handicaps.
The kind of record provided by the orb-web-builder is, unfortunately, not a common phenomenon in behavioral
study, and it would be of questionable usefulness to propose similar investigations with other animals. The generalization which seems appropriate is one in terms of functions. The spider and its web compose a relatively clear system, an instance of a complex but measurable biological operation. Successful reconstruction of it would probably be significant as a model for the operation of other behavioral systems of whatever complexity.
References and Notes
1. A. L. Breed, V. D. Levine, D. B. Peakall, P. N. Witt, Behaviour 23, 43 (1964).
2. P. N. Witt and R. Baum, ibid. 16, 309 (1960).
3. P. N. Witt, Die Wirkung von Substanzen auf den Netzbau der Spinne als biologischer Test (Springer, Heidelberg, 1956). A new, enlarged edition of this book is in preparation.
4. C. F. Reed, P. N. Witt, R. L. Jones, Behaviour 25, 98 (1965) .
5. H. M. Peters, P. N. Witt, D. Wolff, Z. Vergleich. Physiol. 32, 29 (1950).
6. C. F. Reed and P. N. Witt, Federation Proc. 23, 104 (1964).
7. H. M. Peters, Z. Morph. Oekol. Tiere 36, 186 (1939).
8. M. Koenig, Z. Tierpsychol. 8, 462 (1951).
9. A. Christiansen, R. Baum, P. N. Witt, J. Pharmacol. Exp. Therap. 136, 31 (1962).
10. H. M. Peters and P. N. Witt, Experientia 5, 161 (1949).
11. P. N. Witt, Helv. Physiol. Pharmacol. Acta 7, 65 (1949).
12. -, Experientia 7, 310 (1951).
13. -, Helv. Physiol. Pharmacol. Acta 12,
327 (1954).
14. R. Schwarz, Arch. Int. Pharmacodyn. 104, 339 (1956).
15. F. Epelbaum, ibid. 106, 275 (1956).
16. P. N. Witt, ibid. 108, 143 (1956).
17. -, Trans. Coll. Physicians Philadelphia
29, 9 (1961).
18. -, L. Brettschneider, A. P. Boris, /.
Pharmacol. Exp. Therap. 132, 183 (1961).
19. P. N. Witt, unpublished data.
20. D. Wolff and U. Hempel, Z. Vergleich. Physiol. 33, 497 (1951).
21. H. P. Reider, Arch. Int. Pharmacodyn. 115, 326 (1958); Psychiat. Neurol. 134, 378 (1957); Conflnia Neurol. 18, 225 (1958).
22. N. A. Bercel, Arch. Gen. Psychiat. 2, 189 (1960).
23. P. N. Witt and R. Weber, Monatsscht. Psychiat. Neurol. 132, 193 (1956).
24. P. N. Witt, in Psychopathology, A Source Book, C. F. Reed, I. E. Alexander, S. S. Tomkins, Eds. (Harvard Univ. Press, Cambridge, 1958).
25. P. N. Witt, Behaviour 4, 172 (1952).
26. M. Jacobi-Kleeman, Z. Vergleich. Physiol. 34, 606 (1953).
27. B. Fruchter, Introduction to Factor Analysis (Van Nostrand, New York, 1954), p. 59.
28. P. N. Witt, C. F. Reed, F. K. Tittel, Nature 201, 150 (1964).
29. P. N. Witt, Perspectives Biol. Med. 8, 475 (1965).
30. -, Pharmacologist 5 (No. 2), 262
(1963).
31. D. B. Peakall, J. Exp. Zool. 156, 345 (1964); Comp. Biochem. Physiol. 12, 465 (1964); ibid., in press.
32. P. N. Witt, Federation Proc. 21, 180 (1962).
33. -, Arch. Environ. Health 7, 4 (1963).
34. W. N. Schoenfeld and S. H. Baron, Science 147, 634 (1965).
35. P. N. Witt and H. Heimann, Monatsschr. Psychiat. Neurol. 129, 123 (1955).
36. P. N. Witt, Pharmacologist 2 (No. 2), 63 (1960).
37. At various stages in the progress of this work support has been received, and is gratefully acknowledged, from Sandoz Pharmaceuticals, the Swiss National Fund, PHS, and NSF.
8