NOTE: This plain text article interpretation has been digitally created by OCR software to estimate the article text, to help both users and search engines find relevant article content. To read the actual article text, view or download the PDF above.
The Orb Web during the Life of
Argiope aurantia (Lucas)
CHARLES F. REED
Department of Psychology
Temple University
Philadelphia, Pennsylvania
PETER N. WITT
MABEL B. SCARBORO
Division of Research
North Carolina Department of Mental Health
Raleigh, North Carolina
Reed, Charles F. (1969). The Orb Web during the Life of Argiope aurantia (Lucas). Developmental Psychobiology,
2(2): 120-129. Changes in siif, quantity, and regularity of detail were followed in the orb webs of Argiope^^mntia
Lucas for 8 laboratory-born femaleSreared under j^Hrolled environmental conditions. Mesh width and the
number of radii decreased steadily from the 3rd month of life to death. Thread length and area of web reached
peaks around the time of the last molt and sexual maturation, and decreased thereafter. Leg length, weight, and
molt, while related to web characteristics, do not account entirely for changes in dimensions. Furthermore, after
several months under similarly controlled conditions, 6 femal||| that had been caught outdoors built uniSpSally
large webs. The characteristic changes of Argiope aurantia webs laboratory appear to reflect not only
physiological maturation and decline, but may be related to early experience with the efficiency of the web as jn
trap.
garden spider orb-web invertebrate innate behavior maturation early exppienceM field
laboratory data
While its general form remains constant, the orb
web of Argiope aurantia Lucas (Levi, 1968)
changes in size and in number of thread elements
throughout the life span of the animal. Some of these
changes appear to be linear and therefore related to
dimensions of the spider’s body and perhaps to the
output of silk; others show single peak periods proba-
bly attributable to maturation of the spider. Super-
imposed upon these even^ is a considerable daily
variation that is not eliminated by controlling feeding,
temperature, light, humidity, and^çc^sibility to
physical structures upon which the webs are built.
Our purpose in this paper is to describe the develop-
ment of webs of Argiope aurantia in the laboratory
and to attempt to relate it to bodily growth (measured
in body weight and leg length), and to the webs of
Received for publication 20 June 1969.
other species of orb-weavers (Wiehle, 1927; Tilquin,
1942; Witt & Baum, 1960). In attempting to supple-
ment these data, captured spiders were placed under
laboratory conditions after the main collection of data
was underway. Some differences that emerge between
the two samples have challenged assumptions we have
made previdiasly regarding the relevance of the early
history of the spider.
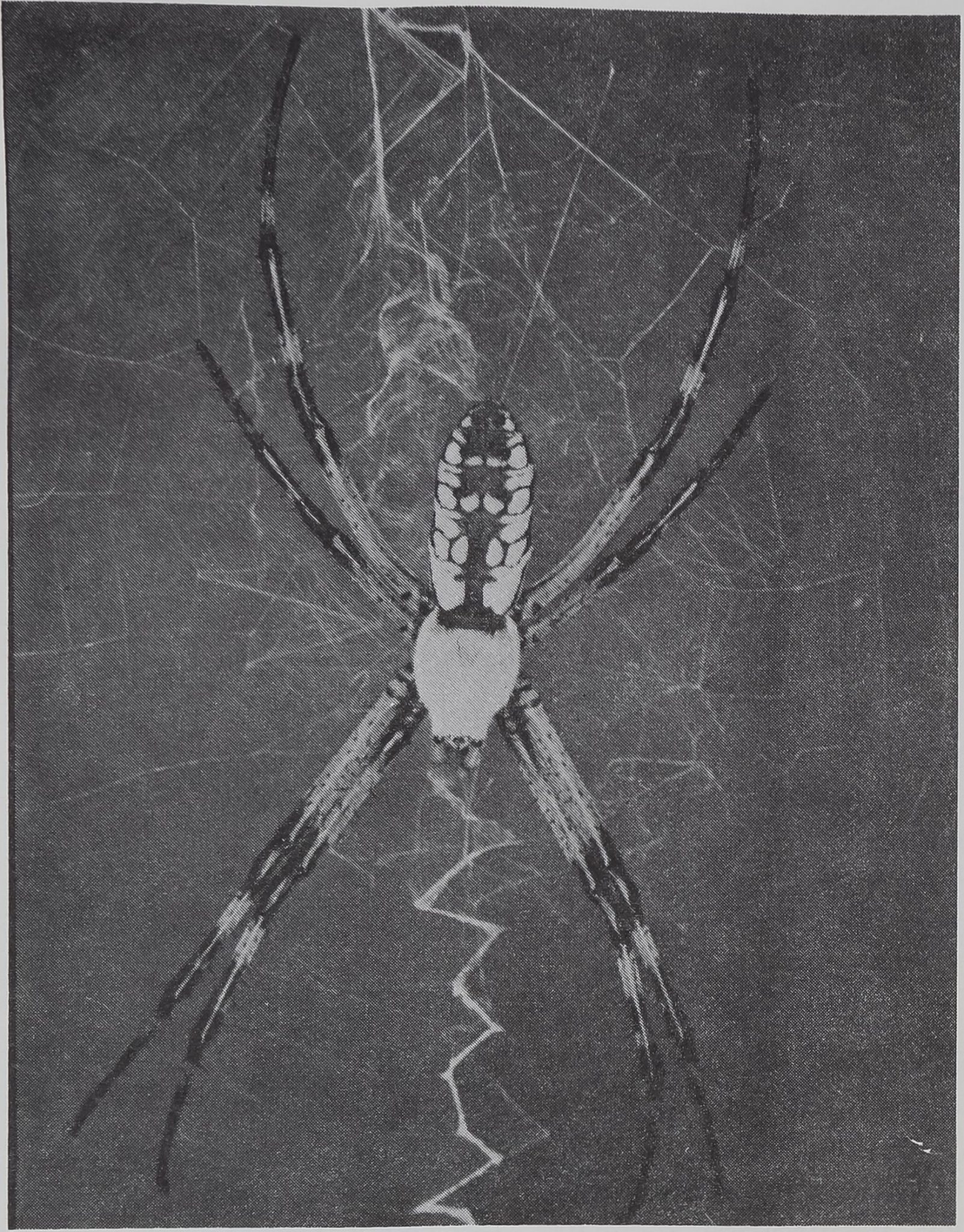
In the laboratory, Argiope aurantia (Fig. 1) molts
five to seven times between hatching and death (12
to 18 months after hatching). There is a close correla-
tion between body weight and length of leg until the
final molt, when the spider is sexually mature and
the abdomen of the female swells with eggs. The
relationship then becomes less clear, but there is pre-
sumably still a complex interaction of organ systems.
Another uncertain period occurs in younger spiders
Developmental Psychobiology, 2(2): 120-129
WEB DEVELOPMENT IN GOLDEN GARDEN SPIDERS
121
fig. 1. This inverted position is taken at the center of the web
by the adult female Argiope aurantia Lucas as it waits for prey.
during molting. Webs built then are small and have
relatively few threads. At molt, apart from the effort
of sloughing the old exoskeleton, neural and sensory
systems are in complex transitional states.
The web of Argiope aurantia is shown in Figure 2.
It is built in the laboratory essentially in a vertical
plane and consists of dry radii supporting the viscid
spiral thread. A hub or platform is separated from
the spirals by a free zone. The hub of the web may
or may not include a stabilimentum: a thick band of
threads placed over the hub of the web at the very end
of construction (appearing as a broad white structure
in the center of Fig. 2). It is not certain that this
structure has a function; none has been identified
(see Discussion). In fact, there is no orderly pattern
of appearance or growth of the stabilimentum through-
out the lifetime of a laboratory-reared spider, as far
as we have been able to determine from examination
of successive webs of individual spiders.
Webs are built fairly regularly on a daily basis,
although there may be periods of several days between
webs. The movements in building webs are con-
siderably slower than those of Araneus diadematus, a
comparatively short-legged species of orb weavers.
METHOD
Eight female spiders were selected from a group
that hatched in mid-January in the laboratory; they
emerged from a single cocoon found outdoors. The
cocoon had been placed in a box filled with gnats,
and the spiderlings could build webs and catch prey
freely. The first molts were observed on January 27
and the first webs on January 29,
At the beginning of March, the animals were moved
to 12 ^16 cm wooden frames, which were in turn
placed in 50 x 50 x 8 cm aluminum and glass cages.
Light was turned on at 8:00 a.m. and off at midnight.
Onset of light was accompanied by a steep rise in
temperature. Hlumidity w^kept at a constant high
level. Each animal received 5 houseflies (Musca do-
mestica) and 5 drops of water per week during this
period.
At the end of March, the wooden frames were re-
moved from the cages, and web-building in the glass
and screen cages was recorded daily. All spiders
continu® on the same feeding regime and were
weighed about every 3 weeks. Mean weight gain for
all animals is shown in Figure 3; individual growth
curves differed considerably; on the same day in May
one spider weighed 108 mg while its equally healthy
litter mate weighed 18.7 mg. We were unable to obtain
any reduction in this variability. The 8 spiders upon
which we report were selected because of their high
production of webs and the full record thereby pro-
vided. Of the 43 original hatchlings, only 22 were
femalS 9 had died by June 1, 12 by the middle of
July, 14 by November.
For purposes of evaluation of webs, the cage doors
were removed every morning, the animal removed
from the web, and the threads sprayed with Krylon
white enamel (development of photographic technique
ultimately enabled us to obtain webs suitable for mea-
suring without spraying). Photographs were taken on
high contrast copy film with the web placed against
a deep black background with fluorescent illumination
from 4 sides.
Points of thread intersection and attachment were
measured in the manner described by Witt et aL (1968).
A computer program extracted a variety of expressions
of web geometry and dimensions from these raw
measures; in general, they dealt with size, regularity,
and shape of the structure. Size measures are the total
length of thread composing the web and the areas of
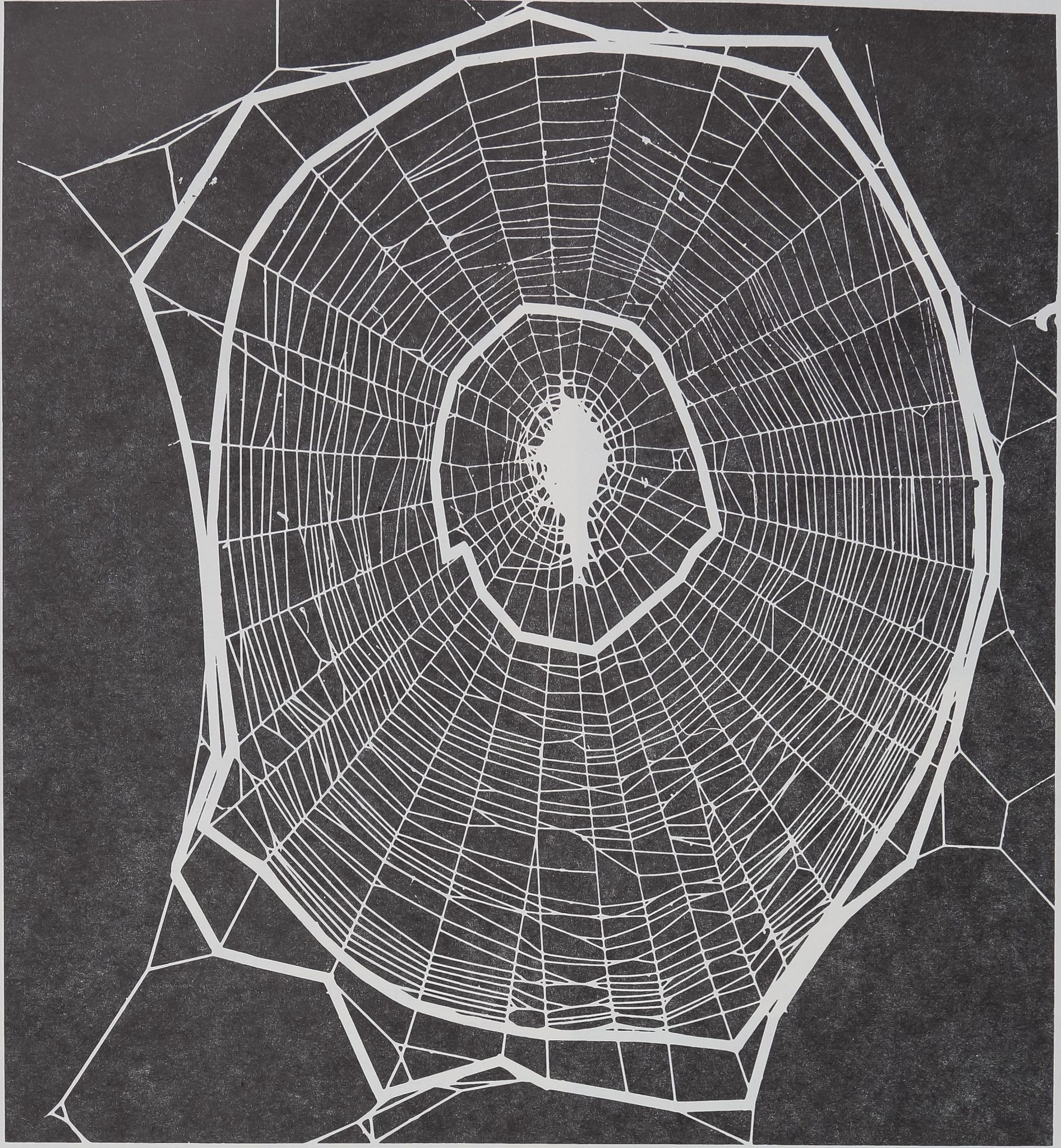
the principal regions of the web. Figure 4 diagrams
these regions; the roughly ring-shaped catching or
spiral area is surrounded by the open spaces of the
frame and scaffolding and encloses the center area
where the spider awaits prey. Regularity refers to the
spacing of threads between the spirals (4 cardinal
122
REED, WITT, AND SCARBORO
fig. 2. Web of adult female Argiope aurantia of captured group, built in October. All avail-
able data indicate that this spider was about the same age as the spider that built the web in
Fig. §f|&). Charact^p%?cally for the whole group, this web is larger and was built with more
radii and spiral turns, Hit has a similar mesh width to the webf%n 5′(&H The weights of the
animals were 177 and 192 mg, respectively, and can hardly explain the difference in webs.
directions were measured), and to the equality
of central angles between radial threads. Shape of the
web is indicated by deviation from a circular form and
asymmetry of the hub with respect to the spiral zone.
Finally, thread density in the catching zone, a web-
index that does not fall into any of the 3 categories,
is calculated by dividing the area of the catching zone
by the product of spiral turns and radial number. This
index, mesh size, assumes that the catching zone is sub-
divided into compartments of equal size (which it is
not); it provides an expression of the expenditure of
thread used to subdivide space. A large catching zone
with many threads and a small catching zone with
few threads can yield identical mesh sizes.
In addition to data for spiders that spent all of their
lives in the laboratory from cocoon to death, we have
obtained web measures for 6 female Argiope aurantia
caught outdoors at the end of July and beginning of
August. Their weights in mid-August corresponded to
the April weights of the laboratory-reared group; we
have concluded from this and other evidence (date
of last molt, death date) that they hatched about 4
months later than the laboratory animals (i.e., at the
usual date for wild Argiope aurantia in the Raleigh,
N. C., area).
After the captured spiders were brought into the
laboratory, both groups spent their lives in identical
cages with identical feeding and environmental
control.
WEB DEVELOPMENT IN GOLDEN GARDEN SPIDERS
123
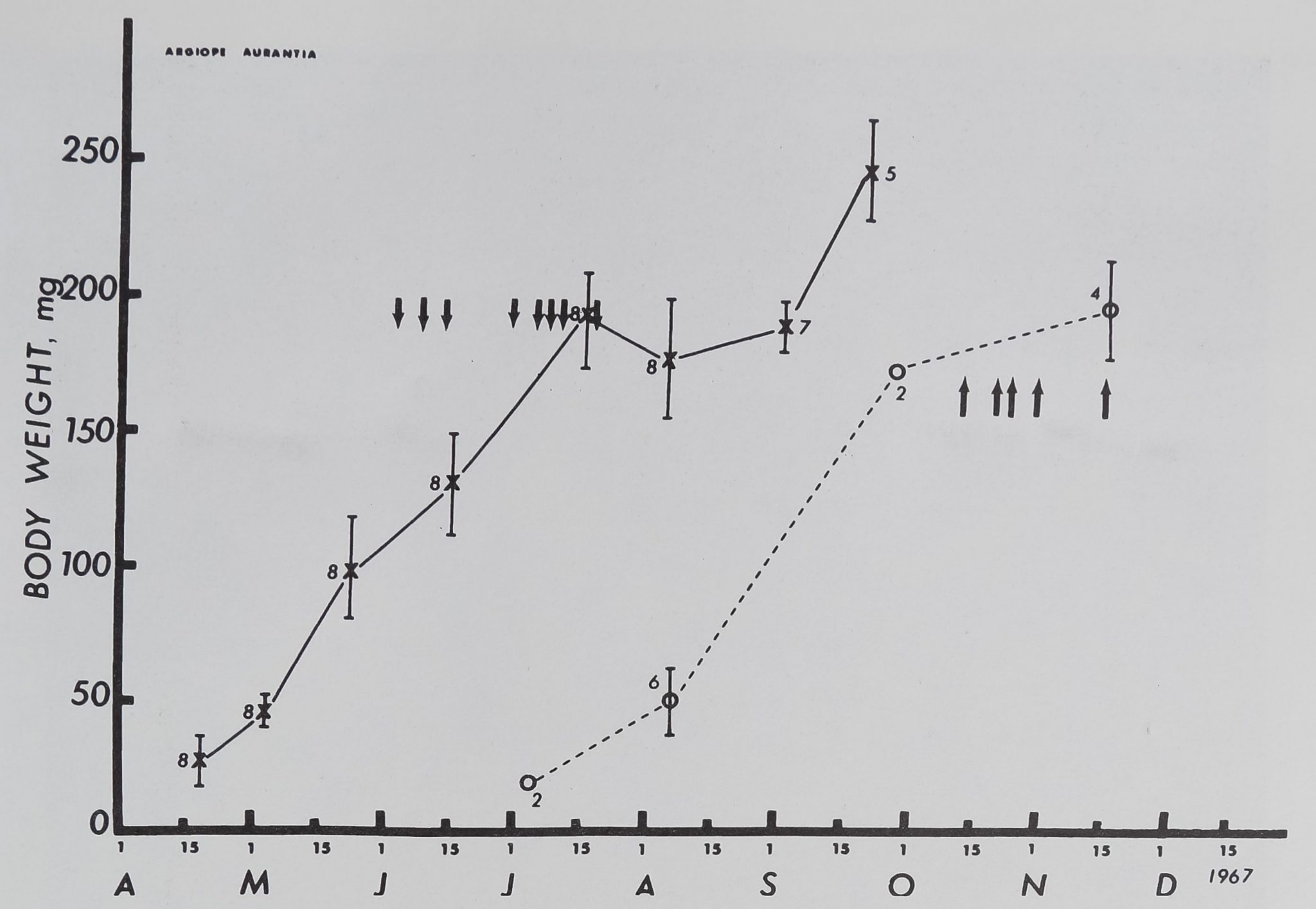
fig. 3. Mean weights (cross£lt and standard errors (line^of female Argiope aurantia littermates
between April 19 and September 25. The mean length of forelegs of the same group in May-mine
was 20.25 ± 0.3 mm. Open circles indicate mean of the few weights obtained for 6 female Argiope
aurantia caught outside; their mean leg length in October 1967 was 23.40 riffi.O mm. Arrows
pointing down show dates on which spiders of the first group molted, arrows pointing up indi-
cat|r> molting-dates for second group.
If one assumes that the unknown hatehifidate of the second group was about 4 months later
than that of the first group, body measures show, in contrast to web measures, no noticable difference
between the 2 groups at comparable ages.
RESULTS
LABORATORY HATCHLINGS
^Blie findings are summarized in Figures 3 to 9. As
Figure 3 indicates, body weight of the laboratory-
born animals increased in an essentially linear manner
until the passing of the last molt. Although not re-
corded on the figure, leg length would show saltatory
rather than continuous growth. Length of the first
leg actually was measured on the molted limb.
It may be noted parenthetically that the chart shows
a similar course for the spiders captured out-of-doors,
although measurements were taken less frequently than
for the laboratory spiders. The 2 curves appear to be
parallel, but displaced in time by about 4 months.
WEB-DIMENSIONS
In the figures that follow, data for all laboratory
animals are combined for each period. This procedure
seemed justified by the approximately equal levels of
building activity. All webs are included in the graphs,
including those built directly before and after molting.
We assumed that the mean accurately represents the
individual, that is, that we are dealing with a single
set of similar curves. The wide variation between
consecutive webs for single spiders makes it very diffi-
cult to che^k^hsftMsumption.
Characteristic webs at 3 life stages for the same spider
are shown in Figure 5.
SIZE AND DENSITY
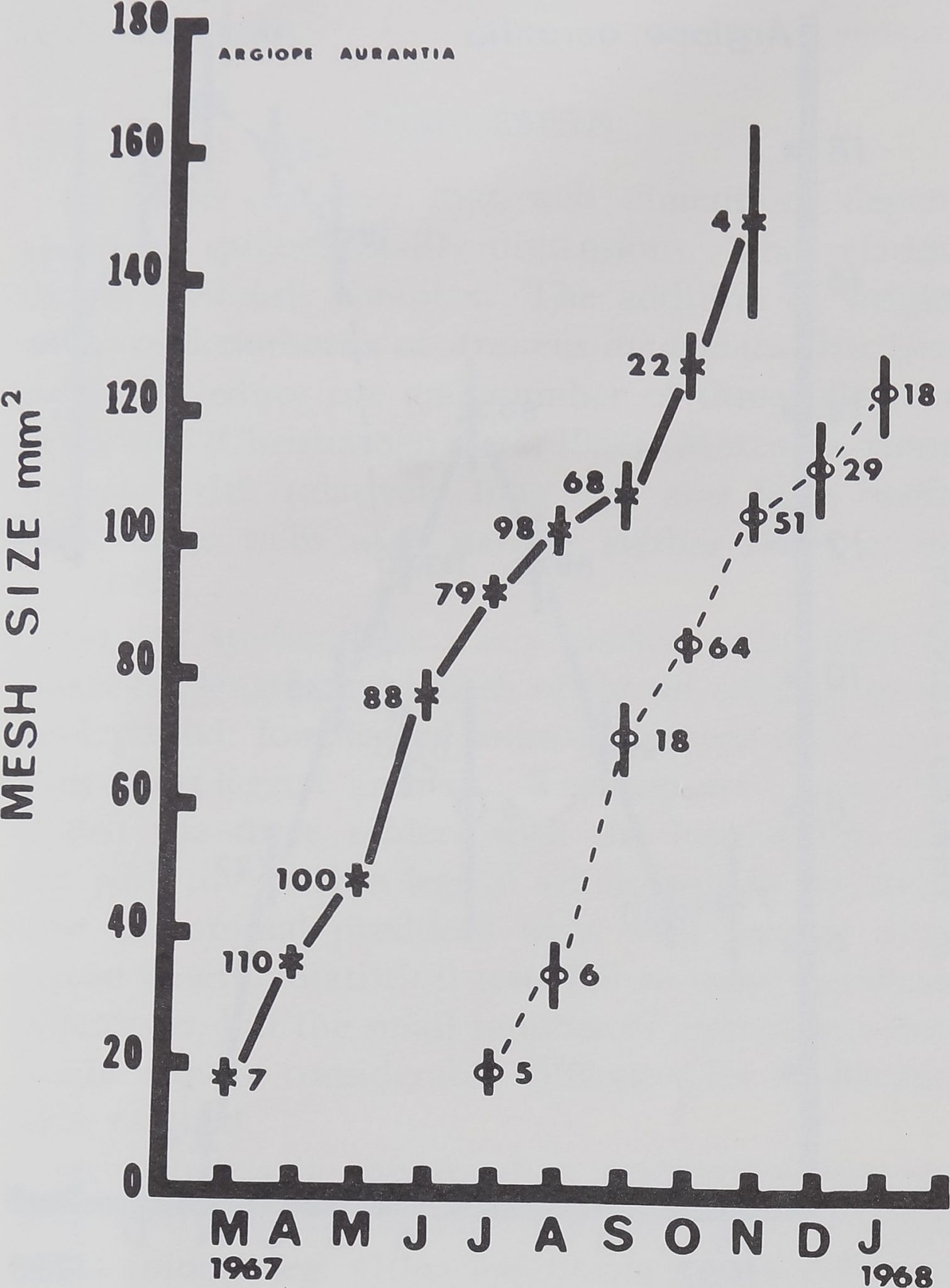
The most orderly course of development occurs in
the most abstract index, mesh size. Figure 6 shows
an Ijsentially linear growth in mesh size throughout
the lifetime of the sample. Moreover, in contrast to
most other measures» the variation in mesh size is
small.Bj|he course of growth is parallel for the labora-
tory-hatchlings and for the captured animals. Since
this measure is an expression of the relative density
of threads in the catching zone of the web, it appears
that as the animal ages the web network assumes a
relatively more open character.
One element in the computation of the mesh size
is the number of radii composing the scaffolding of
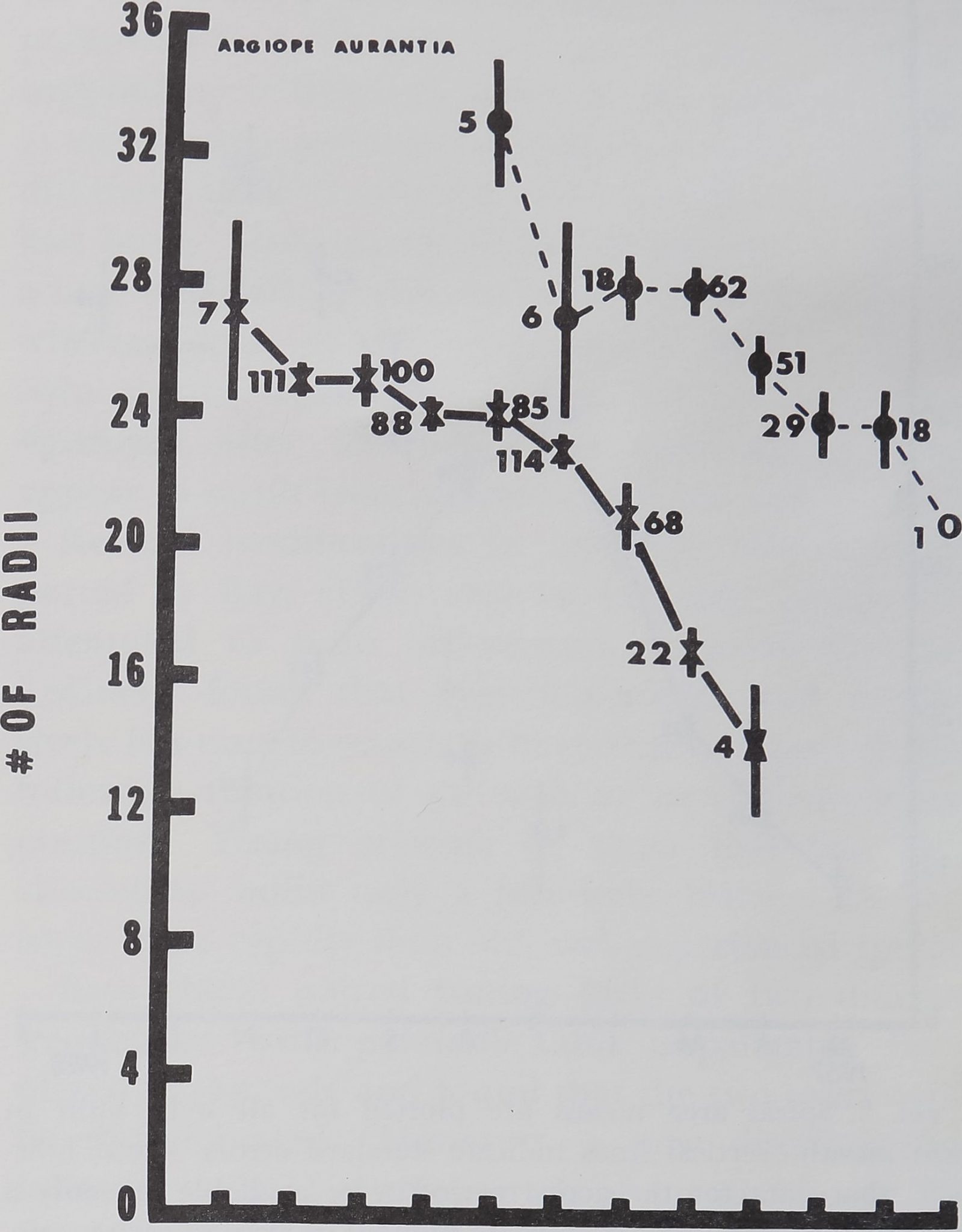
the web. Figure 7 indicates a steady decline in number
of radii for Argiope aurantia, progressively fewer radii
being constructed as the animals age. Tilquin (1942)
124 REED, WITT, AND SCÀRBORO
Fl6‘ iajlBSjSii <Hweb of adult, laboratory-reared female Argiope auramâia. The thick white
lines surround the central area, spiral or catching zone, and frame zone. Note that thS)utermost line
does nil follow the framing threads exactly; this simplification is for computational convMgSjBB
found a relatively small number of radii in the very
first and early webs of Argiope aurantia (Page 81,
Table 3). Our graphs do not include such early webs.
Wiehle (1927) counted higher numbers of radii in the
webs of young Araneus diadematus than in those built
by old animals. The second expression in the denomi-
nator in the formula for mesh size is the number of
spiral turns in the catching zone. These measures
taken separately for the upper and lower part of the
web (not illustrated) range uncertainly in the early
stages of growth, but show a clear decline after July;
a parallel decline occurs for the captured spiders in
November. On the other hand, the numerator in
computation of mesh size, the area covered by spiral
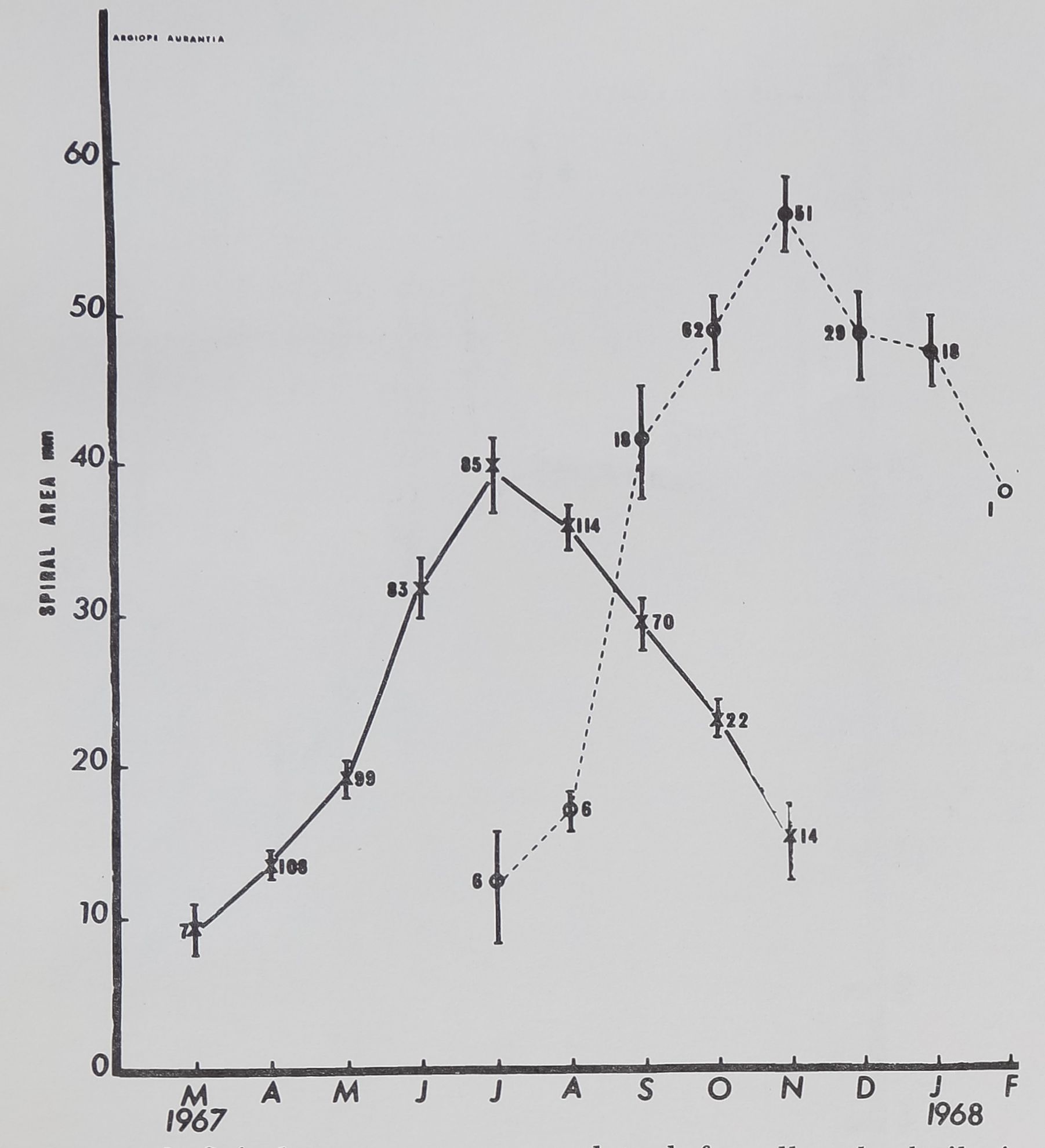
zone (Fig. 8), shows growth and decline. The peak
magnitude for the area of the catching zone is reached
in the July measures for the laboratory hatchlings and
in November for the captured animals.
.-^pThere is thus a kind of reciprocal relationship
between the three expressions that compose the mesh
size measure (area of the zone, number of radii, num-
ber of spiral turns), suchfthat their combined net effect
is to produce a steady reduction in thread density.
Unfortunately, this expression is a statistical abstrac-
tion of uncertain biological significance. It represents
a computed space measure; a quadrilateral space that
would represent an optimal distribution of threads in
the space composing the catching zone. The actual
meshes of the web (the spaces bounded by 2 adjacent
radii and 2 adjacent spiral turns) vary systematically
WEB DEVELOPMENT IN GOLDEN GARDEN SPIDERS
125
A
fig. 5. Three webs built by the same spider (laboratory
group). Webs are shown to scale and were built (a) in April,
body weight 25 mg, leg length 10 mm; (6) in July, body weight
180 mg, leg length 23 mm; and (c) in October, body weight 250
mg, leg length still 23 mm. Note characteristic changes in web
size and detail during 7 months of observation. Mm Figs. 6 to 9.
C
B
from central to peripheral regions of the zone and
unsystematically according to the regularity of place-
ment of radial and spiral threads Fig. 2).
It cannot be inferred simply that the mesh size
matches the capacity’of the spider to span space with
its legsffSi that it is appropriate to the size of the
prey with which the spider is capable of coping. For
the present, we may merely note thatThe density of
thread elements in the web of Argiope aurantia
diminish® throughout the lifetime of the animal.
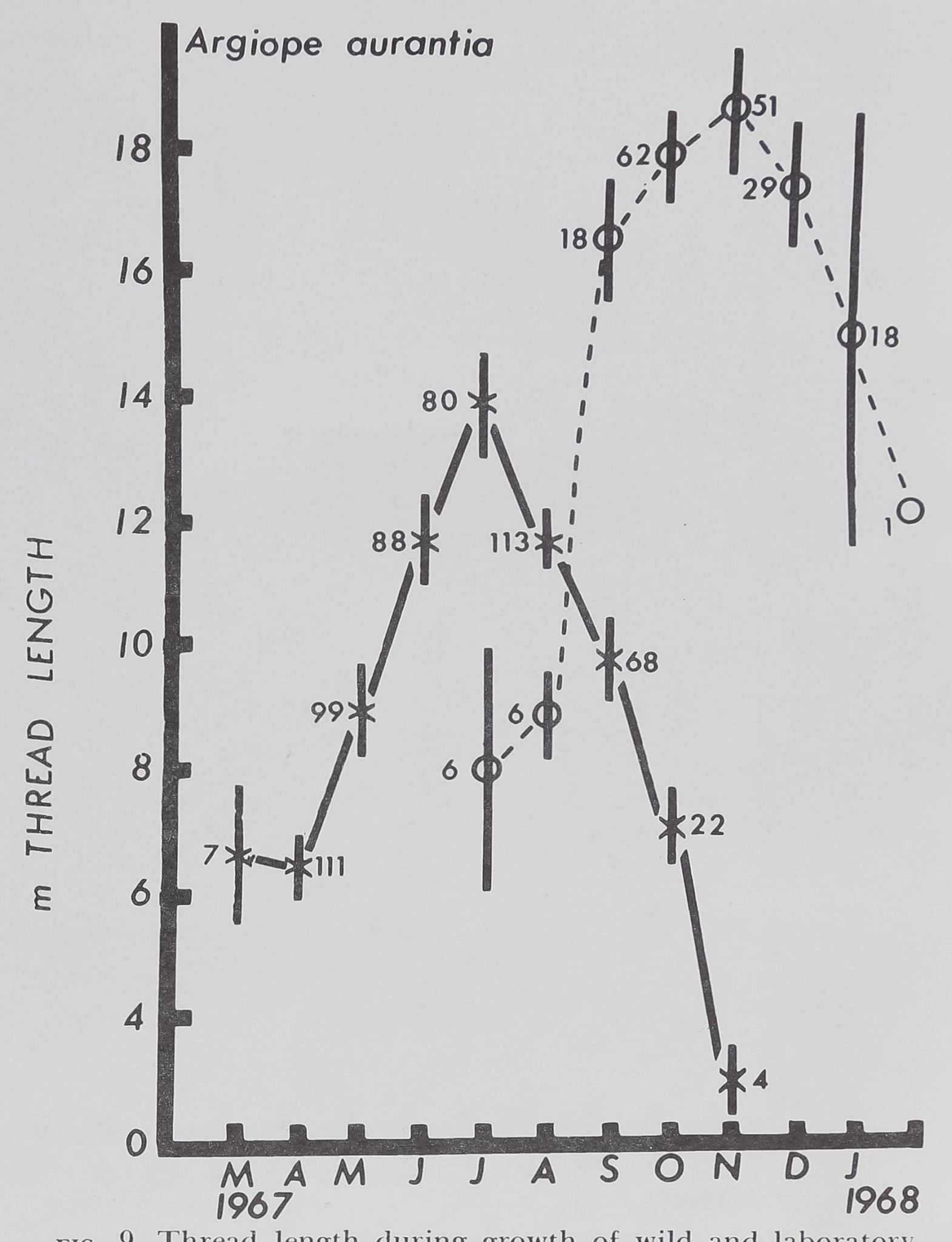
Figure 9 shows that thread length, the total linear
measure of thread used in all parts of the web, di-
minishes during the later part of life. Thread expen-
diture reaches a peak in July for the laboratory hatch-
lings and 4 months later for the captives. Perhaps a
more precise statement would be that the path the
spider takes in building becomes shorter. We have
good reason to believe, but have not proved for this
species, that with increasing weight of the builder the
thread becomes thicket. An old spider’s web, with
relatively short total thread length, could contain as
much material as a web with a relatively long total
thread length built by a lighter (younger) spider
(Christiansen et al., 1962).
The first innermost turn of the sticky spiral marks
the outer boundary of a zone called the center area.
126
REED, WITT, AND SCARBORO
fig. 6. Mesh width (total area ofj^atching zone divided by
number of radii and number of spiral turns) increased evenly
throughout lifetime of both groups.
It includes the hub and free zone. Peters (1937) has
described for Araneus diadematus what is in effect a
positive correlation between leg length and size of
center area. By contrast, our Argiope data show a
peak curve for the center area, similar to that of the
spiral area.
REGULARITY
The foregoing data are concerned with thread out-
put and area of the web. But the web is also charac-
terized by a certain degree of precision of placement of
threads. Adjacent central angles differ little from
each other, the variation of differences is affected by
certain kinds of treatment; for example, removal of 2
ipsilateral front legs severely affects central angle
regularity (Reed et al., 1965). The comparative irregu-
larity of central angle in the older spider could be an
artifact of our computation, which is contaminated by
the relatively larger central angles. The measure for
spacing of spiral turns is, however, relatively precise
and stable.* As shown by an index of relative spiral
deviation, the regularity of this spacing progressively
MA MJJASONDJ
1968
fig. 7. Number of radii, showing decrease throughout ob-
servation period indicated on abscissa. Webs of the captured
group always showed a larger number than did webs of the
laboratory hatchlings.
declines throughout the spider’s life (early webs:
X = 0.46 mm; late webs: X=0.70 mm).
SHAPE AND SPECIAL CHARACTERISTiIIZL
Finally, the overall shape of the web seems to remain
constant; a slightly oval web with hub in the upper
portion remains characteristic.
We have not been able to verify the impression that
young spiders build stabilimenta more frequently
(Comstock, 1948, p. 204) or more elaborately (Levi,
1968, p. 338) than the adult. The stabilimentum was
not given up in captivity, as Wiehle (1927) found
for Argiope bruennichi.
The stabilimenta may differ in role and develop-
ment for species that build them. Definition of the
structure differs with each author. We include the
portion of thread overlaying the hub; other writers
may refer only to the bands extending into the free
zone. Whatever the definition, however, the structure
is highly variable in form throughout the development
WEB DEVELOPMENT IN GOLDEN GARDEN SPIDERS
127
fig. 8. Spiral area means are plotted for all webs built in
each month; fptlod linês indicate standard errors. Note, how-
ever, that data for the^^be period for only 8
spiders. The small figures indicate number of webs in each group.
Note that peak size of zone coincides roughly to time of last molt
and sexual maturation in each group, and is significantly higher
for the wild group, even though t||gy had been in; the laboratory
for more than 4 months,
of Argiope Wjurantia; there is no obvious pattern or
change in complexity.
This variability fails to suppoSany of the functions
hypothesized for the stabilimentum: that it stabilizes
or strengthens the web, conceals or camouflages the
spider, acts as guide or clue for the male’s location of
the female (Crome & Crome, 1961). In the absencë&bf a
contemporary function, it may be possible, j&sVhas
been conjectured by Gertsch (1949, pp. 180-181), that
construction of the stabilimentum is a vestige of once
functional behavior.
A COMPARISON OF BUILDING SEQUENCES
Witt and Baum (1960) have reported web develop-
ment characteristics for Araneus diadematus Cl. and
Neoscona vertebrata McCook. Both species showed
linear functions for body growth (weight and leg
length) and an accompanying linear function for mesh
area. Neither species appears to demonstrate the
peaking effect for spiral area that occurs for Argiope
aurantia Lucas. The number of radial threads at
stages of development may distinguish the 3 species
(very early Argiope webs were not recorded). Number
fig. 9. Thread length during growth of wild and laboratory-
reared Argiope aurantia. Vertical lines indicate standard error
of jPjpte parallel increaseand djüp^^ in curve, with
significant higher values: at peak for captured spiders, follow-
ing peak of laboratory:?group by about 4 months.
of radii for A%giopeT$jp@yf§ a steady decline, Nmkcona
retains a relatively constant number, and Araneus
reaches a plateau after a rapid but negatively acceler-
ated rise in the first 2 months.
CAPTIVES AND LABORATORY HATCHLINGS: DEVELOPMENT
AND MODIFICATIONi|t>F BUILDING-BEHAVIOR
Although the developmental curves for the labora-
tory hatchlings resemble those of the captives, some
differences are apparent.
Several indices of web size show roughly parallel
course^, for the 2 groups of animals, but with peak
levels that are significantly higher for the captives
than for the controls. Figures 7, 8, and 9 contain the
findings for radial number, spiral area, and total thread
length, respectively. These measures are relatively
independent:,,|but conjointly indicate a high level of
activity and expenditure of thread. It is important to
note that development is parallel until the peak period.
This early similarity makes it less likely that selection
REED, WITT, AND SCARBORO
has eliminated those wild spiders that might have
built small webs.
DISCUSSION
We have supposed that web dimensions depend
upon the spider’s bodily dimensions. The relation-
ship is obviously complex. The addition of weights
to the céphalothorax of Araheus diadematus has been
shown to reduce size and number of thread-elemen^
in the web (Christiansen et al., 1962). Moreover, young
Araneus with relatively long legs and light bodies
build large webs with narrow meshes (Witt et al.,
pp. 48-50).
For our spider there was a positive association be-
tween leg length and length of thread only up to the
peak period; long-legged animals covered more space
than short-legged animals. ®he captured group in-
cluded the three spiders with the longest legs and
one with the shortest legs of all animals in the study.
The last animal produced webs with greatest mean
thread length. Statistical tests fail to show significant
differences, but the small number of animals involved
would require considerable difference for significance
to be reached.
Nevertheless it appears that, contrary to our sur-
mise, web size cannot simply be explained by length
of the spider’s leg. Other hypotheses for the difference
in web characteristics seem to be required.
If body dimensions are not sufficient predictors of
web dimensions, what factors might account for the
observed differences? Environmental physical con-
straints were the same for laboratory and wild spiders,
and both groups of spiders had lived in the laboratory
for several months. Several possibilities exist. Sea-
sonal factors may have overridden the uniformity of
laboratory conditions. Alternatively, the first four
months in the wild may have initiated conditions that
became evident only at maturity in the captured
spiders.
With respect to the first possibility, it is true that
the captured animals hatched and grew to maturity
at the time of year characteristic for their species,
whereas the laboratory-reared animals were, in effect,
forced into early hatching.
The possible effect of an early existence in the wild
is difficult to identify. It might be imagined, how-
ever, that the demands for expenditure of thread in
nature are greater than in the laboratory, and that
rate of growth for glands was set by depletion rates
occurring early in the animal’s life. There is no evi-
dence at hand to support this speculation, and it is
not clear how to use it to account for the decline in
thread expenditure that occurs in the later half of the
life of Argiope aurantia.
There is some evidence that feeding experience in
the web affects production of new webs. Spiders never
permitted to catch prey and to feed in the web, but
supplied with flies and water in the same quantities
as web-feeders, produced significantly fewer webs than
did those allowed to remain and feed in the webs they
had built. More particularly, each of the latter built
a new web daily during an 11-day observation period,
whereas none of the other group built every day
(Chi square significance ^f.01). (Webs were always
destroyed after feeding). The reduction does not
appear to result from exhaustion of the web material.
Related modifications in behavior have been re-
ported by Bays (1962) and by LeGuelte (1969), who
attempted to train orb-weavers to particular cues.
LeGuelte found that Zygiella-x-notata took progres-
sively less time to return to its retreat in repeated trials
following rotation of the web to an uncharacteristic
position. Young animals or those that had been
allowed to build only a few webs learned the new
route more rapidly than old, web-experienced spiders.
Bays (1962) paired tuning forks of two different
frequencies with palatable and unpalatable booty
placed in the web, and found that the two tones could
be discriminated. Moreover, a new discrimination
followed reversal of the pairing of cue and booty, and
the learning showed retention over several weeks.
Modification of attack and retreat behaviors appar-
ently is possible for orb-weavers. Modification of the
program of web-building is rather more difficult to
demonstrate. In the first place, there are structural
requirements that cannot be drastically altered. The
web itself is an apparatus for reducing the variability
of the spider’s environment. That is, the spider’s ner-
vous system equips it to deal only with a small number
of contingencies; the demands for learning are few.
However, even in the favorably-biased environment
of the web, there are emergencies and an occasional
need for adaptation (e.g., in catching a potentially
dangerous insect, in repairing damaged portions of the
web). LeGuelte and Bays worked within this limited
repertory of behaviors. The web is a product of a
different form of adaptation than is represented by
learning.
Nevertheless, it is possible that the differences in the
webs of captive and laboratory hatched spiders may be
due to learned behaviors rather than to the glandular
factors mentioned previously. As usual, maturation
must be distinguished from learning. Peters (1969),
for example, has shown that Zygiellae removed from
the cocoon earlier than the normal time for emergence
produce incomplete and irregular webs.
The general question of the possible effects of
experience upon the later history of web construe-
WEB DEVELOPMENT IN GOLDEN GARDEN SPIDERS
129
tion is presently under investigation in this laboratory.
We have in the past assumed, in analogy to computer
operations* that the web is the product of an innate
program that was concurrently modified by input data
regarding environmental and bodily conditions (moor-
ing, weight, thread supply). A possibility that must
now be entertained is that the central program is
susceptible to another kind of alteration required by
the data with which the animal deals early in its life.
Those data could set rates of development for matura-
tion and capacity of organs in the fashion already
hypothesized for the silk glands. On the other hand,
there may be a form of reinforcement of behavior by
the catching of prey that followed web-building. We
have presented some observations suggesting that fre-
quency of building can be affected by the feeding
procedures; perhaps web dimensions are also subject
to thesf’Variables*
NOTES
Supported by National Science Foundation Grant Number
GB-6246X1 to Peterll. Witt.
Mailing address: Charles F. Reed, Department of Psychology,
Temple University, Broad and Montgomery Street, Philadel-
phia, Pennsylvania 19122, U.S.A.
REFERENCES
Bays, S. M. (1962). A study of the training possibilities of
Araneus diadematus Cl. Experientia (Basel), 18: 423.
Christiansen, A., Baum, R., and Witt, P. N. (1962). Changes in
spider webs brought about by mescaline, psilocybin, and an
increase in body weight. J. Pharmacol, exp. Ther., 136: 31-37.
CoM^t’ddK, J. H. (1948). The Spider Book. Ithaca, N. Y.: Corn-
stock.
Crome, W., and Crome, I. (1961M Paarung und Eiablage bei
Argyope Bruennichi (Scopoli) auf Grund von Freilandbeo-
bachtungen an zwei PopMationen im Spreewald / M ark
Brandenburg (Ara^itM AraneidaeWjk Mitt. Zool. Museum
Berlin 37: 189-259.
LEG^Sæ, L. (1969). Learning in spiders. Amer. Zoologist* 9:
*#=452.
Gertsch, W. J. (1949)., American Spiders. Nev^yYork: van
Nostrand,
Levi, H. W. (1968). The spider genera gea and argiope in
America (Araneae: Araneidaew^Bull. Museum Compar. Zool.,
136: 319-352.
Peters, H. M. (1969). Maturing and coordination of web-build-
* mglicfîyity. Amer. Zoologist, 9: 223-228.
Reed, C. F., Witt, P. N., and Jones, R. L. (1965). The measuring
function of the first legs of Aran,eu$ diadematus Clerck.
Behaviour, 25: 98-119.
Tilquin, A. (1942). La toile géométrique des araignées. Paris:
Presses Universitaires de France.
Wiehle, H. (1927). Beitrage zur Kenntnis des Radnetzbaues der
^Éfeiriden, Tetragnathiden und Uloboriden. Zschr. Morphol.
OeMII: Tiêe., 8: 468-53MH
Wm1mBp. |M, and Baum, R. (1960). Changes in orb webs of
spiders during growth (Araneus diadematus Clerck and
Neoscoha vertebrata McCook). Behaviour, 16: 309-318.
Witt, P. N., Reed, C. F., and Peakall, D. B. (1968). A Spider’s
Web. Berlin: Springer Verlag.
Article Title & Publication
The Orb Web during the Life of Argiupe aurantia Developmental Psychobiology