Images Collection


View this article in Search Friendly Plain Text
NOTE: This plain text article interpretation has been digitally created by OCR software to estimate the article text, to help both users and search engines find relevant article content. To read the actual article text, view or download the PDF above.
Physiology & Behavior, Vol. 17, pp. 997—1000. Pergamon Press and Brain Research Publ., 1976. Printed in the U.S.A.
BRIEF COMMUNICATION
Web-Building Time in a Spider: Preliminary
Applications of Ultrasonic Detection
RAYMOND RAMOUSSEM
AND
FRED DAVIS III1 2
Division of Research, North Carolina Department of Mental Health
Raleigh, NC 27611
(Received 24 October 197^H
RAMOUSSE, R. AND F. DAVIS. Web-building time in a spider: preliminary application of ultrasonic detection. PHYSIOL.
BEHAV. 17(6) 997-1000, 1976. – Data collecffln on time and length of building in orb-weaving spiders has suffered from
absence of light during construction and inconVihiint hours. A simpl^fjparatus is described which permits Recording of
the spiders’ movements as they disturb an umÉtee field. By varying onset and, length of dark periods for twqgnimals at
even|f§mperature and by registering the building periods for 127 webs, a definite influence of the light-dark cycle can be
identified: there is a strong preference*Tbr building webs in the dark; this fs superimposed on the circadiaih rhythm of
orhweb^construction. One ^thl fpiders always built earlier than thcoth^^p
Circadian rh^^^S Web-building Onset and length of darkness ^Ultrasol^ detection of movement
SEVERAL MvestigatorlH4, 6, 9] have siudiedmaciters
which influence frequency of orb-web building. The time of
building, however, has only been extensively recorded with
actographic methods in Nemesia caementaria [1] and in
Agelena consociata |£H; these spiders do noTjgJiild orb
webs. In Araneus diadematus or Zygiella-x-notat^th-ere are
only some observations indicating a daip; building time
around 5:30 a.m. |p], or a seasoifi! chkhge frpm 2:00 a.m.
in summer to 5:00|Éf.m. in.tfi^ fall [9]. Light-dark and
temperature cycles have been suspected to play/aj^ole in
setting the time, but the inconvenient hour of building and
the low visibility for direct observation in the early morning
hours have prevented investigators from obtaining sufficient
data.
Knowledge on web-building time is essential for the
understanding of the builder’s survival. An animal like
Araneus diadematus alternates between nearly 23 hours of
immobility, waiting for prey. — only interrupted by short
excursions toward an entangled insect,and abgt one
hour of dismantling the old and constructing the new web.
We can assume that timing is adjusted to maximum chances
of prey catching, and that it varies from species to species
and according to prey activity periods. Does the daily cycle
of building and waiting depend only on the: inner rhythm of
an animal, or can environmental cues cause an adaptation
to specific’-3rcumstances? This preliminary note describes
an ultrasonic methodology and first results of the conse-
quences of light—dark shifts for one orb-web builder, the
crosS-spider Araneus diadematus Cl.
MEfioD
The RWprder
Web-building activity was recorded vtltpc an ultrasonic
system of motion detection. This system was chosen
because:
(1) The threshold response in spiders is observed at
relatively low frequencies, so that the 75.5 kilohertz (kHz)
tone of our instruments is. well beyond the spider’s high
frequency threshold (2,5). It does not seem to interfere
with the spider’s normal activity.
(-2) The wave-length of 75.5 kHz, under normal atmo-
spheric conditions, is 4.39 mm; objects of this size are
easily detectable. The length of the two spiders used was
more than 5 mm.
(3) Movements are permanently recorded on pressure
1 Present address: Laboratoire d’Ethologie Experimentale, 1, rue Pasteur, 69007 Lyon France.
2 Present address: Department of Psychology, Yale University, 10 Hillhouse Av./318, D. L., New Haven, CT 06520.
997
RAMOUSSE AND DAVIS
sensitive paper running at 5 cm/hr. The recorder samples its
output once every 3 sec giving approximately 1200 points
per hr, which is enough to detect any rapid change.
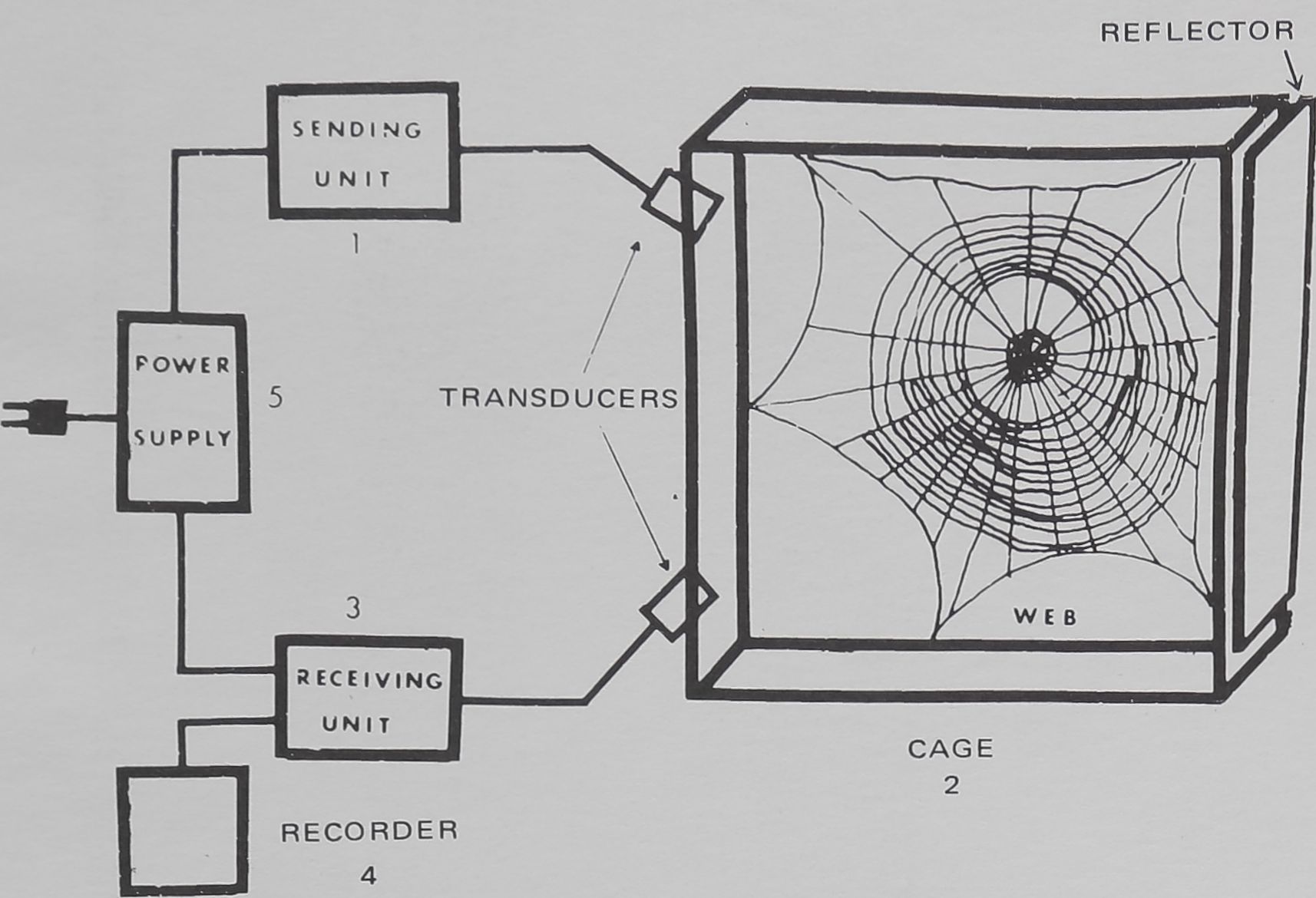
The device (Fig. 1 & 2) consists of five unifs: (1) the
sending unit; (2) The cage; (3) The receiving unit; (4) The
recorder; (5) The power supply of ± 15 and 5 volts for all
circuits.
transducer (which functions as a microphone) and fed via
co-axial cable to the receiving unit. Here they are amplified
to drive the analog recorder.
The National Aeronautics and Space Administration
(NASA) adapted this type of system for the spider
experiments for the Biosatellite program (Witt, personal
communication) but did not use it.
FIG. 1. The five units of the recording device with spider cage and
web.
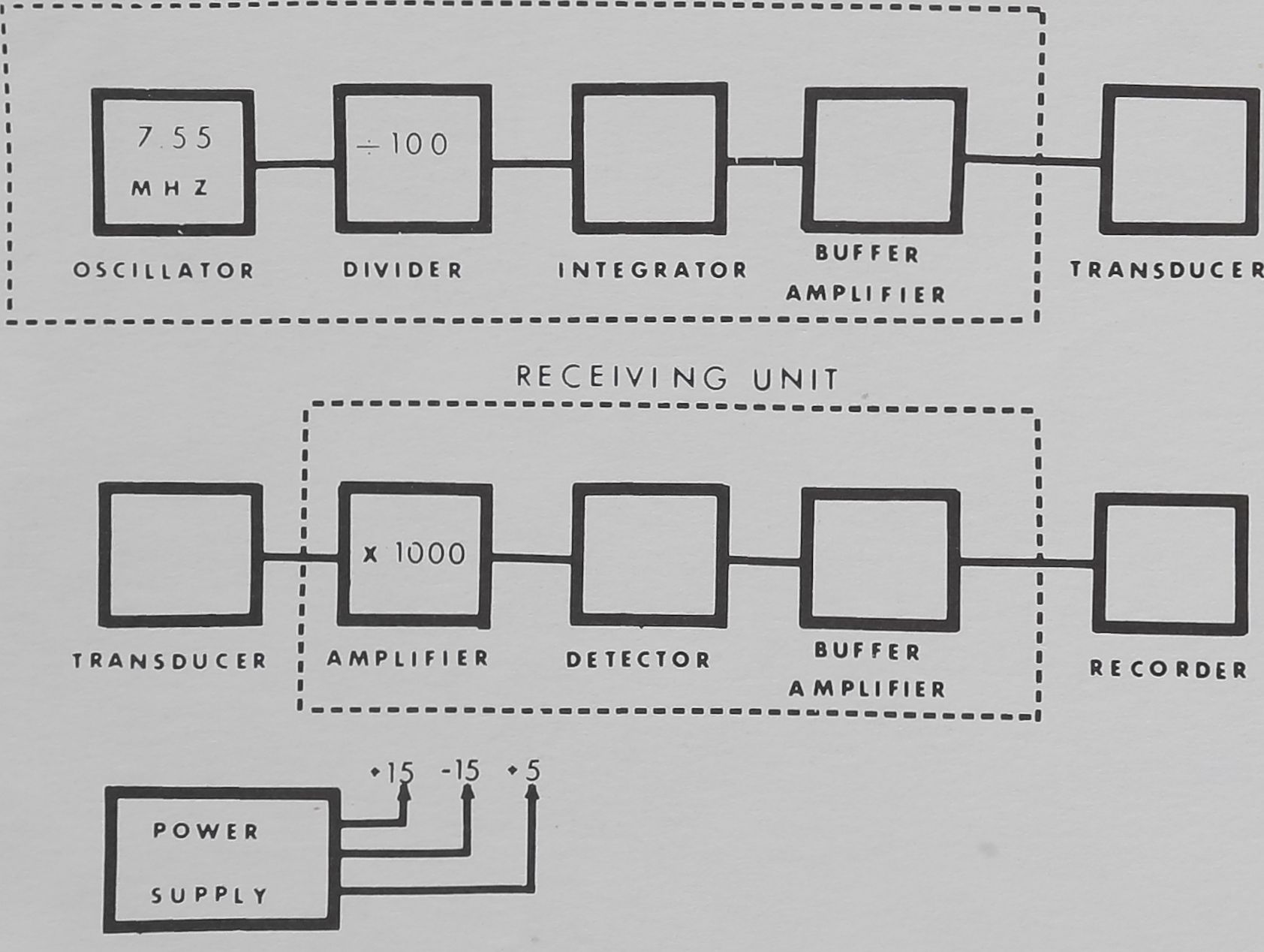
SEND I NG
FIG. 2. Details of the sending unit, the receiving unit and of the
power supply.
The sending unit employs à quartz crystal oscillator for
frequency stability, since shifts in frequency would, produce
spurious signals and drift. The frequency of the quartz
crystal is 7.55 Megahertz (MHz), which is divided to
produce a frequency of 75.5 kHz. The square wave output
of the divider is converted approximately to a sine wave
and fed via co-axial cable to the sending transducer (which
functions as a speaker). This establishes an ultrasonic field
inside the cage.
The cage is a 50 X 50 X 10 cm aluminum frame with
glass sides and screen wire edges. The two transducers are
mounted on one side aimed at the reflector on the other
side. Movements of the spider within the ultrasonic field
create disturbances which are sensed by the receiving
Recording Conditions
Araneus diadematus spiders were placed in* the above
described frames. ThtT*frames were i§; a closed black box
where light and temperature could tie controlled and
recorded (Fig. 3). The box was lit with a 25 W fluorescent
bulb and a 100 W yellow incandescent bulb. Temperature
increased slightly during the light period (Jÿ:C).
HG.ffiMifieriméàffial apparâtu® (1) Black b®l, fron! removed; (2)
Spiders’ cages: one presenting the sound reflector end, the second
presenting the two transduces; (3) The light; (4) The timer; (5) The
sending and receiving units; (6) The linovement recorder; (7) The
temperature recorder.
Two females, A and B, from two different cocoons,
reared in the laboratory (darkness from 0:00 a.m. to 9:00
a.m., 16^; lightly22° C) were studied. Both spiders had
hatched on the same day.
The spiders were fed %. .* housefly and watered twice a
week. As it is known that undisturbed spiders usually spin
their web during the dark period, recording started 3 hr
before the light was turned off and stopped three hours
after the light was turned on. The previous web was
collapsed before the recorder was turned on.
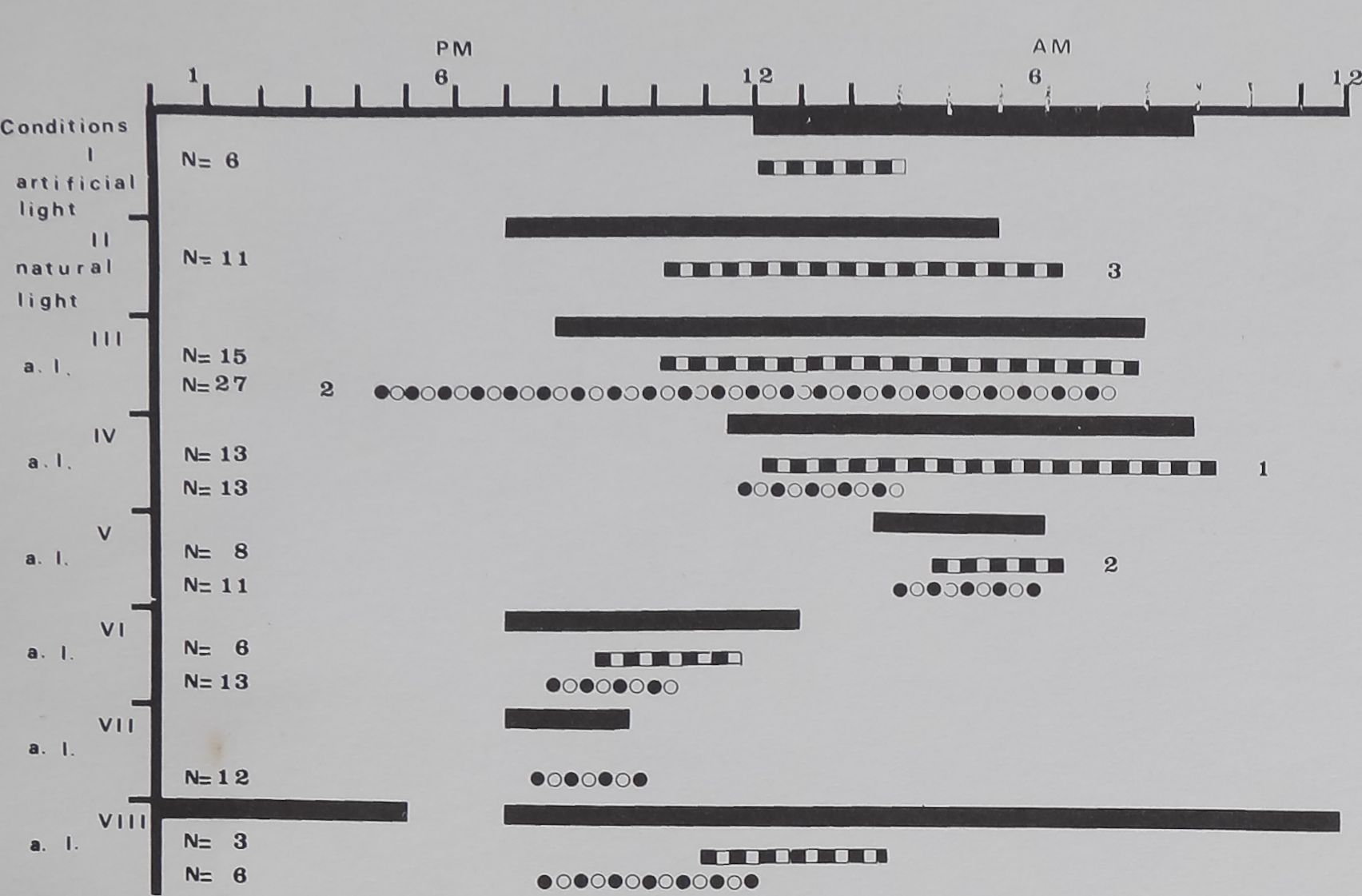
The two spiders were recorded under 8 successive
conditions of varying light (Fig. 4). In the conditions II, VI,
ULTRASONIC DETECTION OF WEB-BUILDING TIME
FIG. 4|- Time of web-building under different conditfeS (from
onset ©f ËÉilding of tb# earliest web to the end H the latest web).
Black bars indicate dark perf®H Black and white£S>H: Spider A,
black and white ciS^ Spider B. l!he number in front IhdigHlthe
numbef of Webs begun before the onH of darkne^srTheSmbers at
the right end indicate the number of webs finished after darkness.
Condition I: condition pf rearing with artificial light. Condition II;
natural light. Condition IIlB VIII: artificial light. Only during
Condition I was there a distiœHemperature <^yêr^^ between
light (= warm) and daifeÉgBoD maintained in the laboratory.
‘ VllBnd VIII onset of darkness y^aM^onstant (7:00 p.m.)
ÿàgnd the length of darkness was M;weegl2 and 22 hr. In
anotheijfset ^^^S>erimenTs” OnHwif darkness was deljàygd
and length of betwSn 3.5 anè ‘%% hr.
A nalvtmmj^R mmrds
Each” i;ecor^hows th#7Q|Hactivity of the -spidejljuring
the recording peri (St In an attempt to differentiate
web-building ^dvity fromHther Ij^RMôfion on a recdgffl
we watehied the rtpordiiH foriBlpr nights. The reJmfe’r
Bad thejppider’s box w£re In^v-ojp^^^’nt rooms in orde§|
not; to disturb the Bcordmg by Rn^lder movement^He
txperimente^^Jked at the spider in th^ramefech time a
signal appeared ©Hthe record® During IslSIM b||sts of
^ activity (Ipsa than 5 min) we always saw the spider
exploring. In iB ftase Hi long burst w^aw wenp^g
and. exploratory activity. The signals’ during web-building
were always larger in Amplitude than during explorations
and web-bugteg Exploratory
phase (Fig. 5). ExploratioiF’^^greplacé the period
formally:^contains destruction Hp^lMd webfend whicM
usually nr Mëdés building of the Jrew structure.
RESULTS AND DI^ÿsi<pJ|
Duration of Web-Building
The mean lengths ,of web acjiyify were: Spider A M
80.295. 22.01 min, N = 78 ^ebHSpider bB7$J4 -±^18.6|
min, N = 49 webs. No significant differences exist between
the two means.
The duration of web-building activity, in a night is
positively correlated with size of spiral area built in that
night (Kendall rank correlation, A and B, p – 0.001),
indicating that it takes more time to build a larger web.
Building Time
The two spiders started to build their webs always in
999
FIG. 5. Sample read downward: (A) Record of web-
-building onlyH(B) Reo(Él|Hs? webffiilding and. exploration.
We^giMingjisKhe last activity?,recorded, and the magnitude of the
signals of web-building is larger than the^s^afs of explosion; (C)
Record with iHBivity at all.
exceptions which ©furred near the
molting fimfl^of Spider B (Condition III), Sometimes
building «atted clcÉlgito the flHof darkness and was
finisHd ^uMng the light period. Most of the time the
spidftjl bdH a web during KB figt pew ëfark period
Blowing the shift from one Bndition to the other.
When onset of ;darknes$Jkaried the delay between onset
%f |||rkneH and onset ©I web-Mlding decreased with
Increasing length of darkness Hriedmah two-way analysis
of variance, Spiders A and B, p = 0.01). Web-building took
place later aft||§ onset of darknlll when darkness was
longer.
These reslIH suggest that Araneus diadematus builds
webs preferably in the dark, where onset of darkness may
be a positive (releasing) factor of web-building and onset of
lighwp hegative (inhibitory) one. When the dark period
lasted very long (Condition VIII), web-building appeared to
follow the inner clock more closely. There was also a
distinct difference between Spiders A and B, the latter
always building somewhat earlier.
According to these observations the dark-light cycle
definitely influences web-building even if there is no change
in temperature; but only observations of many spiders over
1000
RAMOUSSE AND DAVIS
long periods of time can establish a more exact relationship
between the circadian rhythm of web-building and roïë of
darkness. It also remains to be seen in the future whether
different spiders have individual timing, as they have
individual web patterns [I The new method described
here can establish such correlations1 2 3 4 5. Web construction at
night can be understood as an efficient device which
protects the builder from visually oriented predators, it also
provides the spider with the full light period for- catching
airborne prey. Spiders which specialize on piéy which is
active at night may show other construction tim^fe
ACKNOWLEDGEMENTS
t This work was carried out in the laboratories of the Division of
Research, North Carolina Department of Mental Health and was
suf plpei by Grant Number GB 25274 from the National Science
Foundation to Dr. Peter N. Witt. The. authors gratefully acknowl-
edge the assistance of Dr. Witt during all stages and the assistance of
Mrs. Mab^Scarb’oro-Tor all technical and laboratory work and Mrs.
Rubenia Daniels for her administrative assistance.
REFERENCES
1. Buchli, H. Le rythme d’activité <chez^la^^gale maçonrig*
Nemesia caementarià (Ctenizidae). Am. feÿiiwyfïe’s 19:.
195-196, 1968.
2. Finck, F. Vibration^^ptlvity^ an orb-we,âVSî5 AMuZé^i. 12:
539-545, 1972.
3. Krafft, B. Various aspects of tj^biology of Agelerig, e®$sociata
Denis when bred in the labof|§Ofy. Am. Zool. 9: 201—210,
1969.
4. Peters, H. M. Uber das Kreuzspinnennetz und seine Problème.
Naturwissenschaften 47: 776—786,1939.
5. Walcott, L. and W. G. van der Kloot. Th©-physiology of the;
spider vibration réçe^toï.,/. exp. Zool 41: 19ilp®|jja^59.
6. Witt, Pr?N- ïDièffflirkung fort Subspahzen âuf den Netzbm dèê
Spinne ah biologischer Test. Berlin-Go ttingen-Heidelb erg :
Springer^ 1956.
7. Witt, P. N. Environment in relation to the behavior: of spiders.
71-1.2,1963. I
8. Wife P. Égj.’ J. O. Rawlings and C. F. Reed. Ontogeny of
web-building behtvibf in two oi^^^aving spiders. Am. Zool.
12: 445-454, 1972.
9. Wolff, D. and U. Hempel. Versuphe ;über die Beeinflussung des
Netzbaues von Zilla-x-notata durch Pervitin, Scopolamin und
St^hnin. Z. vergl. Physiol. 33: 497-®8,1951.